INTRODUCTION
Toxoplasma gondii is an obligate intracellular protozoan parasite which causes congenital defects or abortion and fatal diseases in immune compromised patients. As many as 15-85% of individuals in adult human population show serological evidence of T. gondii infection world-widely (Jackson and Hutchison, 1989). The ingestion of tissue cysts in undercooked meat from intermediate hosts or the ingestion of food or water contaminated with oocysts from feces of definitive hosts are the two major modes of transmission (Dubey et al., 1997). Experimental transmission of T. gondii by certain Felidae (Jewell et al., 1972) and high prevalence of antibody against T. gondii in animals of Felidae (Choi et al., 1987) supported the possible candidates of these animals as the final hosts of T. gondii indirectly. But only cat has been confirmed as unique definitive host (Frenkel et al., 1970) that can excrete the environmentally resistant oocysts until now.
Pet cats recently infected may shed large numbers of oocysts in the household, thereby putting their owners at risk from the infection (Pereira et al., 1992). Cats allowed to roam free or stray cats may contaminate the environment with oocysts which may infect human or livestock (Tenter et al., 1994). Therefore, cats play a key role in the epidemiology of toxoplasmosis and the availability of a reference test for the detection of T. gondii infection in cats are needed. We demonstrate here the seroprevalence of stray cats by western blot analysis against T. gondii, comparison of western blot with ELISA and indirect latex agglutination test (ILAT), and the diagnostic availability of sandwich-ELISA with monoclonal antibodies.
MATERIALS AND METHODS
Antigen preparation
RH tachyzoites of T. gondii were maintained by a peritoneal passage in inbreeding mice (ICR strain) and purified by centrifugation over 40% Percoll (Amersham Pharmacia Biotech, Uppsala, Sweden) in PBS solution (Choi et al., 1992).
Sera
A total of 198 sera from stray cats was collected from 1996 to 1998 at suburban and rural areas near Chinju City, Gyeongsangnam-do and was freezed at -70℃ until use. Sex and age of cats were not identified.
Western blot, ELISA and ILAT
Western blot was done by the method of Towbin et al. (1979). The whole RH extracts were separated in 12% SDS-PAGE gels and transferred onto nitrocellulose (NC, Schlleicher and Shuell, Keene, NH, USA) papers. NC papers were incubated with sera of 1:200 dilution, and then with 1:5,000 diluted HRP-conjugated goat anti-cat IgG antibody (Jackson ImmunoResearch Lab, West Grove, PA, USA). They were soaked in enhanced chemiluminescence (ECL) solution (Amersham Pharmacia Biotech) for 1 min and exposed to an X-ray film (Konica, Tokyo, Japan) for 5 to 10 sec.
ELISA was performed according to the method described previously (Choi et al., 1992) except the serum dilution of 1:500 and HRP-conjugate dilution of 1:10,000. ILAT was done by the method described in a kit manual (Eiken Toxo-kit, Tokyo, Japan) and titers of 1:32 or higher were regarded as positives.
Sandwich-ELISA with monoclonal antibodies
For the selection of monoclonal antibodies (mAbs) to be used in sandwich-ELISA, mAbs were confirmed by immunofluorescence assay (IFA) and western blot. Freshly prepared RH tachyzoites cytospinned onto 18-mm coverslips and Vero cells (CRL 6318, American Type Culture Collection, Rockville, MD, USA) cultured on coverslips infected with RH tachyzoites for 24 hr were fixed either with cold absolute methanol for 5 min or with paraformaldehyde for 10 min and then permeabilized with Triton X-100 for 5 min, separately. mAbs were applied with 1:100 dilution in incubation solution (3% BSA in PBS) and coverslips were incubated with 1:500 diluted FITC-conjugated goat anti-mouse IgG antibody (Sigma Chem Co., St. Louis, MO, USA). Fluorescence was observed under a fluorescence microscopy (Axiophot, Carl Zeiss Co., Oberkochen, Germany).
Sandwich-ELISA was done as follows. Subcellular organelle (surface membrane, dense granule, rhoptry, and cytosolic) specific mAbs were coated onto a 96-well EIA plate (Costar, Dover, NH, USA) with 1:100 dilution in coating buffer (50 mM carbonate buffer, pH 9.6) overnight at 4℃. After blocking with 5% skim milk in PBS/0.05% Tween-20 (PBS/T) for 1 hr, RH tachyzoite extracts of 200 µl were incubated with 10 µg/ml in blocking solution for 2 hr. After then conventional ELISA procedure was adapted as described above except serum dilution of 1:200 and HRP-conjugate dilution of 1:10,000.
RESULTS
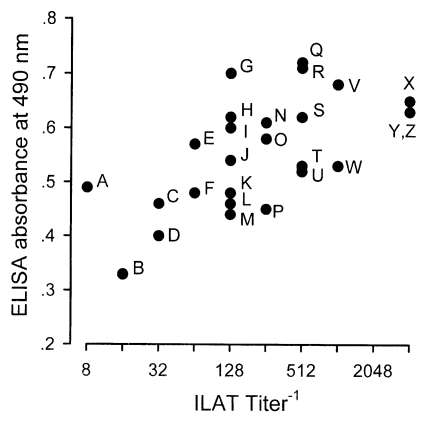
Typical western blot patterns against T. gondii were found in 26 sera (13.1%) out of 198 stray cat sera. The patterns revealed a set of major surface protein reacting bands with molecular weights of 30 kDa (SAG1, Mineo et al., 1993) and 22 kDa (SAG2, Parmley et al., 1992), 22-30 kDa dense granular proteins (Cesbron-Delauw, 1994), and 50-70 kDa rhoptry proteins (Sam-Yellowe, 1996) with some minor bands (Fig. 1). When spotted by ILAT titer and ELISA absorbance as shown in Fig. 2, 24 cases (92.3%) were in the positive range of 1:32 or higher and 2 cases (lane A and B) in reactive but negative range by ILAT, and all 26 cases were distributed over cut-off value of 0.275 calculated as mean+3 SD of 124 ILAT-not-reactive cases (mean, 0.140l; SD, 0.045) by ELISA.
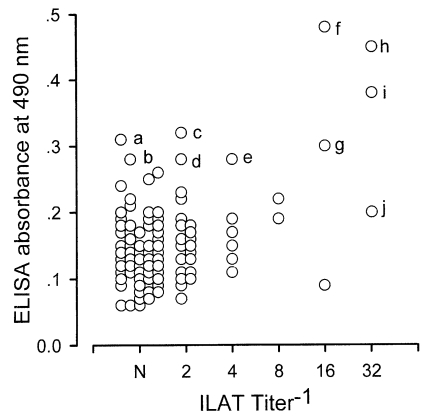
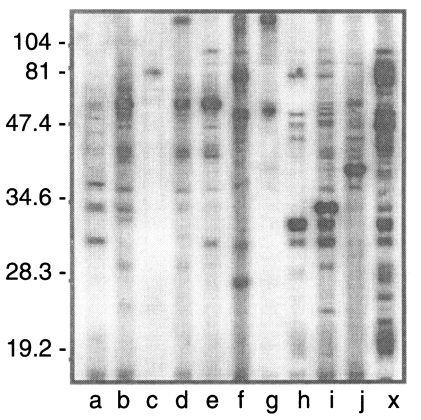
Western blot negative 172 sera were distributed as shown in Fig. 3. Three cases (h, i, and j) were ILAT positive (1:32) and 9 cases (a to i) were ELISA positive (over 0.275) with 2 cases positive by both ILAT and ELISA (h and i). Western blot of these 10 cases were presented in Fig. 4 with one positive serum (lane X) in addition. Each serum showed various binding patterns with RH antigens but not typical as described above.
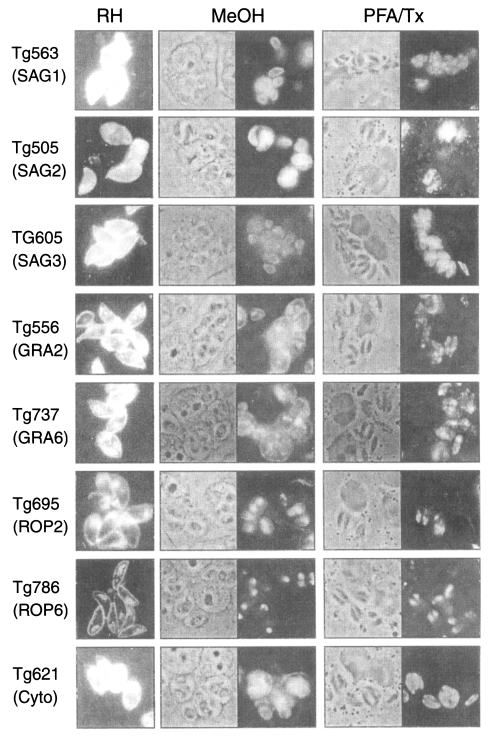
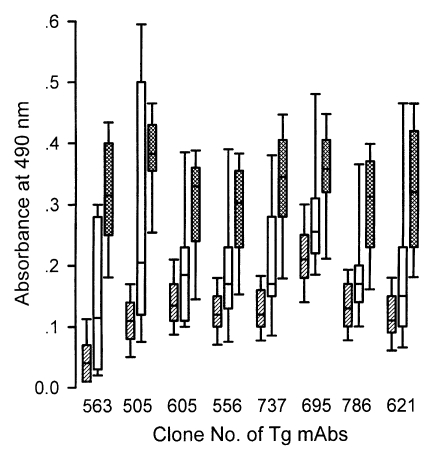
To clarify the reactivity of these sera in western blot, ELISA or ILAT, we performed sandwich-ELISA with mAbs which were confirmed by western blot (Fig. 5) and IFA (Fig. 6). Tg563 clone detected 30 kDa protein in western blot and surface membrane in extracellular RH (designated by RH in Fig. 5) and both in methanol fixed intracellular RH (designated by MeOH) and PFA/Tx, which represented SAG1. Tg505 and Tg605 clones also showed similar IFA patterns with Tg563 clone except for positive reaction with 22 kDa protein (SAG2) and 43 kDa (SAG3) by western blot, respectively. Both Tg556 and Tg737 clones labelled extracellular and intracellular granules with parasitophorous vacuolar membrane (PVM), but immunoblotting detection of Tg556 was 28 kDa (GRA2) and that of Tg737 was 32 kDa (GRA6), respectively. Tg695 and Tg786 clones labelled rhoptry proteins of 66 kDa ROP2 and 42 kDa ROP6. And Tg621 clone labelled unidentified cytosolic 32 kDa protein which was secreted into PVM.
By the sandwich ELISA with mAbs (Fig. 7), 124 ILAT-not-reactive cases were distributed as negative and all 26 western blot-positive cases were in the positive range clearly with cut-off value calculated by the absorbance using each mAb. Among the 10 false reactive cases, 7 cases were revealed to be in the negative range but 3 cases (lane a, h, and i) were in the positive range with one or more mAbs showing high absorbance as summarized in Table 1.
DISCUSSION
A total of 26 cases (13.1%) out of 198 stray cats was detected to be seropositive against T. gondii by western blot. This low seropositive rate was comparable to the European and American regions with high seroprevalence rate approximately over 40% in cats (Frenkel et al., 1995; Hill et al., 1998). Since cats as the final host of T. gondii are related directly with T. gondii transmission to human by shedding of oocysts (Choi et al., 1997), low seroprevalence rate among cats in this study may reflect low seroprevalence rate of T. gondii antibodies in Korean people of 2 to 10% according to the group tested (Choi et al, 1989, 1992). The prevalence of T. gondii infection varies according to the life style of cats such that it is higher in feral cats than those in domestic cats (Dubey, 1973; Nogami et al., 1998). But pet cats are exposed to be free-ranging stage easily, it is suggested that pet cats are not much less infected with T. gondii than stray cats in Korea.
The western blot patterns of T. gondii infection has been established well in such reactions with major surface membrane proteins, dense granular proteins, and rhoptry proteins. Therefore, it is a method of choice to detect antibodies in the sera of infected individuals. All 26 cases of western blot positive were detected as positive in ELISA, but 2 cases were missed by ILAT (Fig. 2). However, out of 172 western blot negative sera, 9 cases were in the range of positive by ELISA and 3 cases were positive ILAT titer of 1:32 (two cases were positive by both ELISA and ILAT). Western blot of these 10 cases revealed peculiar patterns not comparable to each other. Irregular reactivities might be resulted from non-specific bindings or the epitopic similarity of T. gondii antigens with other pathogenic infections common to cats such as Isospora felis (Omata et al., 1991), Hammondia sp. (Hendricks et al., 1979), and feline immunodeficiency virus (FIV) and feline leukemia virus (FeLV) antigens (Svobodova et al., 1998) especially.
Indeed, mAbs used in sandwich ELISA seemed to have equivalent sensitivity to the conventional ELISA using specific antigens purified or prepared by recombinant expression of p30 (Lunden et al., 1993; Nam et al., 1996), p22 (Parmley et al., 1992), and GRA4 (Mevelec et al., 1992). All mAbs specifically labelled subcellular organelles each other and clearly discriminated negatives from positives as depicted in Fig. 7. When applied to the 10 false reactive sera by western blot, 7 cases were defined as negative and 3 cases were still positive (Table 1). In case of lane a of Fig. 4, only Tg506 clone reacted in sandwich-ELISA, which suggested the serum could detect p30 with several other bands. And in cases of h and i of Fig. 4, all the mAbs reacted strongly despite they did not react in western blot clearly. Therefore, it is suggested to be elicited during the course of infection because there are no differences in binding patterns between surface membrane proteins and subcellular organelles.
All mAbs used in this study need further investigations to be more specific to T. gondii infection as a standardized sandwich-ELISA to differentiate it from other pathogens.