INTRODUCTION
Toxoplasma gondii is an obligatory intracellular parasite that causes severe disease after congenital infection and in immunocompromized patients such as AIDS. During infection, immune effector cells are able to kill or inhibit its intracellular growth. This antiprotozoan activity produces a number of toxic products such as reactive oxygen intermediates. And within the host cell, T. gondii itself produces oxidants as by-products of normal metabolism.
Reactive oxygen species (ROS) are potentially destructive, capable of oxidizing proteins or lipids and causing chemical modification to nucleic acids (Halliwell and Gutteridge, 1991), even though some of these species also function as second messenger molecules (Dalton, 1999). Superoxide dismutase (SOD), glutathione peroxidase (GPx), and catalase are the three major enzymes to be involved in the cellular anti-ROS system. Hydrogen peroxide, which is produced by SOD at the initial step of ROS detoxification, is subsequently eliminated by catalase and/or Gpx (Halliwell and Gutteridge, 1991). The presence of endogeneous SOD, GPx, and catalase have been demonstrated in T. gondii (Hughes et al., 1989; Ding et al., 2000; Odberg-Ferragut et al., 2000; Kaasch and Joiner, 2000), which suggests to be related with protection from ROS attacks.
Peroxiredoxin (Prx) is a recently described family of antioxidants that are highly conserved in eukaryotes and prokaryotes (McGonigle et al., 1998), which has been called as thioredoxin peroxidase (Tpx,) or thiolspecific antioxidant (TSA). Prx acts as an antioxidant enzyme by eliminating hydrogen peroxide and hydroxyl radicals. The catalytic mechanism of the enzyme involves the redoxactive cysteine, which is highly conserved in the vicinity of the 47th position of its amino acid sequence (Chae et al., 1994). We report here the cDNA cloning, expression, and functional characterization of a Prx from T. gondii (TgPrx).
MATERIALS AND METHODS
Parasite
The RH strain of T. gondii was maintained by peritoneal passages in Balb/c mice. Tachyzoites were purified by centrifugation over 40% Percoll (Amersham Pharmacia Biotech, Uppsala, Sweden) in PBS solution (Sohn and Nam, 1999).
Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR) with redundant primers
Total RNA was purified from the RH extract with Tri Reagent (Sigma Chem. Co., St. Lous, MO) and used as template. RT-PCR was performed with the redundant primer set (forward primer, F1: 5'-AC(A/T/C) TT(C/T) GA(A/G) TG(T/C) CC(T/C) AC(A/G) GA-3' and reverse primer, R1: 5'- TT(G/T/A) GCC GG(A/G) CA(T/C/G) ACT TC(T/A) CC-3') designed near the two active cysteine residues of Prx in other organisms (Yamamoto et al., 1989; Ishii et al., 1993; Chae et al., 1994a; 1994c). Amplified DNA fragment of appoximate 380 bp was cloned into pGEM-T vector (Promega, Madison, WI) and sequenced to design gene specific primers (GSP).
5'- and 3'-Rapid Amplification of cDNA End (RACE)
5'-RACE and 3'-RACE were performed according to the method in Frohman et al. (1988) with primers designed as 5-CDS: 5'-CTA ATC GAC TCA CTA TAG GCA AGC GTG GTA CAA CGC AGA GT-3', 5-NEST: 5'-A AGC GTG GTA CAA CGC AGA GT-3', GSP1: 5'-CAC GTT TCT CCA GGC GTT GTG CAC-3', GSP2: 5'-CAC CGA TTG CAT GGC GAG TTC GAG-3', GSP3: 5'-GTG CAC AAC GCC TGG AGA AAC GTG-3', GSP4: 5'-AGA GCG TCC AAC ATG CGA AGG GCT-3', 3-CDS: 5'-AAC CAG TGG TAA CAA CGC AGA GTA CTT TTT TTT TTT TTT TTT TTT TTT TTT GC-3', and 3-NEST: 5'-AAC CAG TGG TAA CAA CGC AGA GT-3'. Sequences from 5'-RACE and 3'-RACE were combined to be a full-length sequence of TgPrx.
Production of anti-recombinant GST fusion proteins antiserum
RT-PCR was performed to amplify the DNA fragments of open reading frame of TgPrx and superoxide dismutase (TgSOD) as a control. Primer sets used were as follows; for TgPrx, PRXF: 5'-ATG CCG GCC CCG ATG GTG TCT-3' and PRXR: 5'-TTA CTT GCT TCC GAG ATA CTC-3', and for TgSOD, SODF: 5'-ATG GTA TTC ACT TTG CCC CCG-3' and SODR: 5'-TCA TTT CAA GGC ATT CTC CAA-3' based on the gene in GenBank accession No. AF029915 (Odberg-Ferragut et al., 2000). Amplified DNA fragments of TgPrx (591 bp) and TgSOD (606 bp) were cloned in pGEM-T Easy vector (Promega) to subclone in pGEX-4T vector (Amersham Pharmacia Biotech). TgPrx and TgSOD were expressed as GST fusion proteins in HB101 bacteria by the IPTG induction. Polyclonal antibodies against recombinant TgPrx and TgSOD were raised in rabbits immunized and boosted 2 times with glutathione-sepharose purified GST fused TgPrx and TgSOD, respectively. Sera were collected at the 6th week after initial immunization.
Cell culture and artemisinin treatment
Vero cells (CRL 6318, American Type Culture Collection, Rockville, MD) were maintained in DMEM supplemented with 10% FBS (Gibco BRL). Vero cells and Vero infected with T. gondii for 24 hr were incubated in 0.1% FBS/DMEM for 6 hr prior to treatment of 1 µg/ml artemisinin (Berens et al., 1998). The whole extracts were assayed with antisera of TgPrx and TgSOD in addition to a mammalian Prx I (Kang et al., 1998) as a control.
Immunofluorescence assay (IFA)
Tachyzoites from mouse peritoneal passage were attached to 18-mm coverslips by cytospin. Vero cells cultured on 18-mm coverslips in 24-well plates were infected with tachyzoites for 24 hr. IFA was done according to the procedure of Sinai et al. (1997). Tachyzoites were fixed with methanol for 5 min and cells were fixed either with methanol or with 3% paraformaldehyde for 10 min and then permeabilized by 0.05% Triton X-100 for 5 min, separately. Polyclonal anti-TgPrx rabbit antiserum was diluted in 1:100 of 3% BSA/PBS, and mouse monoclonal antibody against dense granular protein (clone No. Tg378 in Son and Nam, 2001) was used as a control for the PVM involvement. Rabbit antibody was detected with TRITC-conjugated goat anti-rabbit IgG antibody and mouse antibody with FITC-conjugated goat antimouse IgG antibody (both Sigma Chem. Co.) in 1:500 dilution. Fluorescence was observed under a fluorescence microscopy (Axiophot, Carl Zeiss Co., Oberkochen, Germany).
Western blot
Western blot was performed by the method of Towbin et al. (1979). The cell extracts were separated on 12% SDS-PAGE gels and transferred onto nitrocellulose sheets (NC, Schlleicher and Shuell, Keene, NH). NC papers blocked by 5% skim milk in PBS/0.05% Tween-20 were incubated with monoclonal antibody for SAG1 (clone No. Tg563 in Son and Nam, 2001) and rabbit antiserum of 1:1,000 dilution, and then with 1:2,000 diluted HRP-conjugated goat anti-mouse and anti-rabbit IgG antibody (Cappel, Costa Mesa, CA). They were soaked in enhanced chemiluminescence (ECL) solution (Intron, Daejon, Korea) for 1 min and exposed to an X-ray film (Konica, Tokyo, Japan).
RESULTS
The DNA fragments of 634 bp (Fig. 1, lane 1) and 744 bp (Fig. 1, lane 3) were amplified by 5'-RACE and 3'-RACE on the cDNA of tachyzoites, respectively. Both the DNA fragments were confirmed to be specific to T. gondii by nested PCR. Nested PCR with 5-NEST and GSP1 primers produced a 424 bp DNA fragment (Fig. 1, lane 2) on the DNA fragment of lane 1 as the template, which made a shorter fragment of 210 bp as expected. And nested PCR with GSP2 and 3-NEST primers amplified a 675 bp DNA fragment (Fig. 1, lane 4) with the DNA fragment of lane 3 as the template which resulted in 70 bp difference. A full-length cDNA sequence of TgPrx was obtained by combining the sequences of the DNA fragments of lane 2 and 4 of Fig. 1. It was 1,101 bp long including a predicted open reading frame of 591 bp. Conserved 2 cysteine domains were present as Phe-Val-Cys-Pro and Glu-Val-Cys-Pro in deduced 196 amino acid sequence (Fig. 2), which indicated that TgPrx belonged to 2-Cys Prx families. The cysteine residue of the anterior domain localized at 51 amino acid (Cys-51) and that of posterior at 171 amino acid (Cys-171). A search of the Toxoplasma database of expressed sequence tags (EST) identified a sequence of 197 bp (Ctoxqual2_665) with perfect match. This EST sequence is overlapped with the anterior cysteine (Cys-51) domain and used in the analysis of peroxidoxins by McGonigle et al. (1998). Deduced amino acid sequence of TgPrx revealed significant sequence similarities with other 2-Cys Prx (Fig. 3).
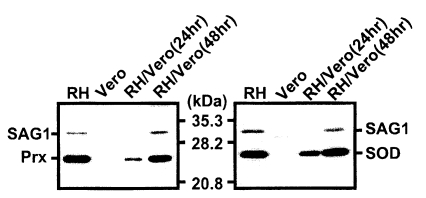
TgPrx and TgSOD were expressed as GST fusion proteins in HB101 bacteria by the IPTG induction with molecular mass of 51 kDa and 52 kDa, respectively (data not shown). Polyclonal antibodies against recombinant TgPrx and TgSOD detected those proteins of T. gondii each other specifically without binding to host cells (Fig. 4). TgPrx was detected as 25 kDa in extracellular and intracellular stages as traced by anti-SAG1 monoclonal antibody and TgSOD as 26 kDa with the same pattern. TgPrx was localized in the cytoplasm of tachyzoites as revealed by IFA images (Fig. 5A) comparing with spot distribution of dense granular protein (Tg378). Its distribution was limited in the tachyzoites and was not secreted into PV or PVM whereas Tg378 was secreted into PVM during the intracellular stages (Fig. 5B).
Artemisinin, a sesquiterpene endoperoxide, was treated to trace the changes in TgPrx after being exposed to the peroxide during the intracellular growing stage of tachyzoites. The concentration of LD50 was 1 µg/ml in our culture system (data not shown). Artemisinin itself evoked little changes in host cell Prx as detected by anti-mammalian Prx I antibody (Fig. 6A). However, the expression of TgPrx increased as early as 30 min and reached to the plateau in 1 hr by the treatment with artemisinin in the intracellular stage (Fig. 6B), while there were little changes in those of host Prx I and TgSOD. Host cell Prx had approximately equal response to artemisinin regardless of infection by T. gondii.
DISCUSSION
A full-length cDNA sequence of TgPrx was composed of 1,101 bp including a predicted open reading frame of 591 bp, which coded 196 amino acid sequence of 25 kDa TgPrx. It was preceded by a T-rich region in the 5' untranslated region which was seen in other T. gondii genes (Cesbron-Delauw et al., 1989; Ossorio et al., 1992; Mercier et al., 1998; Lecordier et al., 1999; Carey et al., 2000). The first ATG did conform to the T. gondii consensus translation initiation sequence of NAAA except for C at position +4 (Seeber, 1997).
Two cysteine residues were positioned at 51 amino acid (Cys-51) and 171 amino acid (Cys-171) as compare to 47 (Cys 47) and 170 (Cys 170) in other Prx (Chae et al., 1994a; 1994b). These site variations were also found in tryparedoxin peroxidase (Alphey, 2000). All the sequences of anterior cysteine domains of Prx compared were the same as Phe-Val-Cys-Pro, and the posterior also as Glu-Val-Cys-Pro except for Entamoeba histolytica of Ala at the first position. The amino acid sequence of TgPrx showed the highest homology with that of Arabidopsis thaliana by 53.9%. Among Prx of parasitic organisms, Prx of E. histolytica had the homologous sequence of 39.5%, which made this organism belong to a cluster in upper hierarchy with AhpC of Escherichia coli (33.1%).
Prx are found in all organisms and their primary sequences are highly conserved. This ubiquity and high degree of conservation suggest a biological importance of this enzyme. Prx are critically involved in cellular defense, receptor signaling, protein phosphorylation transcriptional regulation, and apoptosis (Zhang et al., 1997; Jin and Jeang, 2000). TgPrx was also abundant in tachyzoites as in other eukaryotic cells of which the Prx covered 2 to 5% of total cellular proteins as a housekeeping protein (Chae et al., 1994b). And TgPrx localized in the cytoplasm of tachyzoites and maintained in intracellular stage without secretion into PV nor PVM, which suggests its role in defense mechanism against ROS produced during the metabolism within tachyzoites, although the exact function of TgPrx is still unknown.
Artemisinin, active constituent of qinghaosu as an endoperoxide which has been effective as antimalarial drug (Meshnick et al., 1996; Cumming et al., 1997), inhibited also the intracellular growth of T. gondii in vitro (Berens et al., 1998) by imposing oxidative stress within the cells. It has been proposed that artemisinin activated by the intraparasitic metals catalyzes the cleavage of this drug to ROS (Meshnick, 1998). In addition, ROS can also be cleaved from the drug in the cytoplasm of host cells, which then delivered across the PVM to reach the tachyzoites within the PVM. Resulting ROS might then damage the parasite by alkylating and poisoning one or more essential proteins. Inducible expression of Prx has been demonstrated for many mammalian cells. Inducers includes hydrogen peroxide (Kim et al., 1997), heme/heavy metals (Immenschuh et al., 1997), and others. Changes in TgPrx against artemisinin, in our results, may reduce the ROS by binding with its active thiol group. Therefore, since TgPrx is sensitive to artemisinin, it is suggested to be a good candidate for chemotherapy.