INTRODUCTION
The number of vivax malaria, which was once regarded as eradicated from this country in late 1970s, has rapidly increased every year since 1993 in the northern part of Gyeonggi-do, Republic of Korea (Chai et al., 1994; Cho et al., 1994; Kim et al., 1996; Lim et al., 1996; Shim et al., 1997; Lee et al., 1998; Shim and Kim, 1999; Lee et al., 2000).
Understanding the dispersal patterns of vector mosquitoes in nature is essential to evaluating the risk of malaria transmission to human, implementing surveillance, and developing abatement strategies. It is known that malaria transmission is closely related to the population density and the flight range of the vector mosquitoes (Trpis & Hausermann, 1986). There have been many studies on the females flight range of Anopheline spp. in 1900s. Wright (1918) described the flight range of An. sacharovi to be 13 (Kligler 1932)- 14 km (Kligler & Mer, 1930; Bruce-Chwatt, 1970), based on larval habitat to search overwintering site. Rosenstiel (1947) reported that the flight range of An. freeborni is over 42 km before hibernation. It has also been reported that Culex tarsalis is able to fly at least 17.7 km per night (Bailey et al., 1965) and Aedes taeniorhynchus can fly long distance up to 32 km from breeding sites, when accompanied with wind (Johnson, 1969).
Despite many studies worldwide concerning the flight range of various species, the only study done in Korea has been with Culex tritaeniorhynchus, the main vector of Japanese encephalitis virus (JEV) (Ree et al., 1975).
In this study, we measured the flight range and patterns of Anopheles sinensis, the main vector of malaria in Korea.
MATERIALS AND METHODS
Study areas
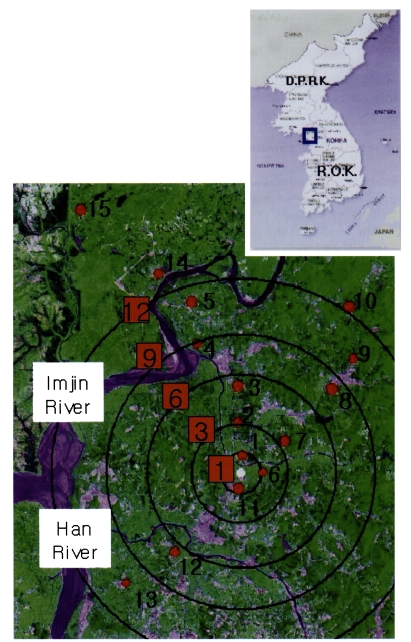
This study was conducted from 7th to 28th September 1998 in Paju city, where there had recently been re-emergence of malaria. The study area is located near the demilitarized zone (DMZ) between South Korea and North Korea. The study area covers rice paddy fields and roads among which small villages was scattered toward south-west (SW) and north-northwest (NNW). Many mountains are located on north-northeast (NNE), and Imjin- and Han-River are located to the north-west (NW). During the study period, the livestock census of the dispersal area were 1,647 cows and 9,483 pigs in Paju city, 1,915 cows and 12,697 pigs in Munsan-eup, 5,533 cows and 9,896 pigs in Beobwon-eup, and 668 cows and 7,110 pigs in Wollong-myeon. Many rice paddy fields and small villages were located within 1 km around the center of dispersal (Fig. 1).
All weather conditions were obtained from Monthly Report of National Institute of Weather, and rainfall, wind speed and wind direction were recorded continuously at the release site.
Collection, mark and release methods
We collected unfed females of An. sinensis with an aspirator from 20:00 to 22:00 at cattle sheds in Paju city. All the mosquitoes were placed into the cages (45 × 45 × 45 cm) and then transported to the releasing site on the same day. Before release, the mosquitoes were marked with two kinds of fluorescent pigments (Sung Kwang Chem. Co., Korea). Twenty five grams of each pigment were diluted with 100 ml ethanol and applied. On the first day, 6,439 female mosquitoes were marked by jetted spray with yellowish fluorescent dye, and on the third day 6,334 female mosquitoes were marked with orange dye. They were all released outdoors in the releasing point (Fig. 1) on the same day.
Recapture
In order to recapture the marked females of An. sinensis, we set up light traps at 3 directions of NNW, NNE, and SW. Ten light traps were set at 1, 3, 6, 9 and 12 km from the release point in directions of NNW and NNE, and only 3 light traps were set at 1, 6 and 9 km from the same point toward SW because appropriate cattle sheds could not be found. Total 13 light traps were operated in this study. Additionally, we also set up two light traps at 16 and 20 km from that point toward NNW to monitor the longest flight range of mosquitoes. These two light traps were operated only 3 times, because the areas are located close to the DMZ and the entrance of civilians is highly restricted.
All light traps were set up outside of the cattle sheds and were operated from 18:00 to 06:00 for 21 successive nights after release. All captured mosquitoes were transported to our laboratory and An. sinensis mosquitoes were selected. To check whether the mosquitoes were marked or not, 32 mosquitoes were placed on each filter paper (150 mm, Advantec, Tokyo, Japan), and one drop of 100% ethanol was dropped onto the body for spreading fluorescent dyes from mosquitoes. The ethanol was evaporated and the filter papers were observed under UV transilluminator.
RESULTS
Weather
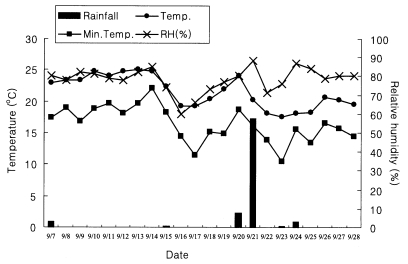
The period of dispersal experiment was from early to middle of September, and the climate of high temperature and humidity was influenced by high atmospheric pressure of North Pacific. The temperature was around upper 20℃ during this time, which was 1-3℃ higher than the annual average, but the number of rainy days and amount of rainfall were lower than usual. Although the wind blew openly toward southeast, wind condition of dispersal areas was not regular. The average velocity of the wind was 3 m/sec (Fig. 2).
Main area of activity and duration of survival
One hundred ninety four female mosquitoes were sorted according to the distance from the release site. 72 female mosquitoes (37.1%) were recaptured within 1 km from the release site, 57 mosquitoes (29.4%) in 1-3 km, 41 mosquitoes (21.1%) in 3-6 km, 20 mosquitoes (10.3%) in 6-9 km, and 4 mosquitoes (2.1%) in 9-12 km. Total 170 female mosquitoes (87.6%) out of 194 marked mosquitoes were recaptured within 6 km. It was therefore concluded that the main area of activity was within 6 km. 45 female mosquitoes (23.2%) were recaptured on the next day after releasing. one mosquito was captured in light trap which was set at 12 km from the release site. Until 20 days after the beginning of release, the released mosquitoes were captured, indicating that An. sinensis survived more than 20 days in the field. About 80% of the marked mosquitoes were recaptured within one week (Table 1).
Longest flight range
To estimate the longest flight range, we had 2 light traps installed at 16 km and 20 km from the release sites (No. 14 and 15 of Fig. 1), which were operated 3 times during this experiment. Total 4 marked female mosquitoes were recaptured. Therefore, the vector of malaria in Korea can fly quite a long distance (data not shown).
Recapture rates
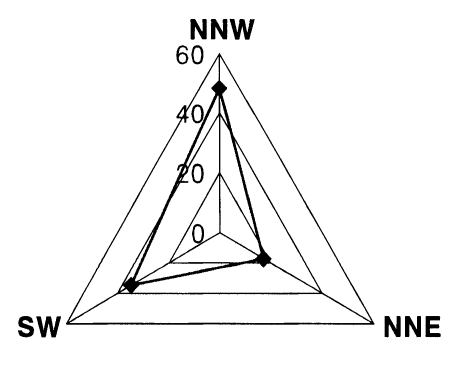
Of the 12,773 released females of An. sinensis marked with fluorescent dyes, 194 female mosquitoes were recaptured by 13 light traps between 7th and 28th September. The recapture rate was 1.52% (194/12,773) (Table 2). In NNW direction, 27,208 female An. sinensis mosquitoes were captured in 5 sites. Among them, 66 female mosquitoes marked by yellow fluorescent dye (n=6,439) were recaptured and the recapture rate was 1.03% (66/6,439) for this color. And 48 female mosquitoes marked by orange fluorescent dye (n=6,334) were recaptured, so the recapture rate of this color was 0.76% (48/6,334). In this direction, 114 female mosquitoes were recaptured, so the recapture rate was 0.89% (114/12,773). The number of recaptured mosquitoes in this direction was much more than those of another two directions; that is, 114 female mosquitoes from total 194 female mosquitoes recaptured (58.8%) (Table 3).
In NNE direction, 8,975 female mosquitoes of An. sinensis were captured on 5 sites, and 17 female mosquitoes were marked by yellow and 22 female mosquitoes by orange color. So for this direction, the recapture rate was 0.31% (39/12,773). Compared with other two directions, the recapture rate was relatively low 20.1% (39/194) (Table 4).
In SW direction, 19,809 female mosquitoes of An. sinensis were captured on 5 sites, and 41 female mosquitoes marked by fluorescent dye were recaptured. Among them, 15 female mosquitoes were marked by yellow and 26 female mosquitoes by orange color. So the recapture rate was 0.32% (41/12,773) (Table 5).
Main direction of flight
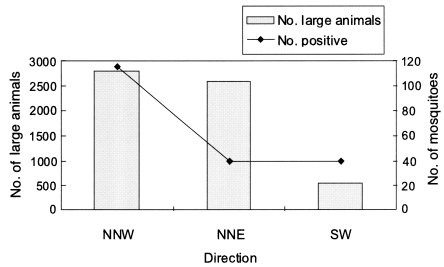
According to their recapture rate, the main direction of flight of An. sinensis in this experiment was shown to be 58.8%, 20.1% and 21.1% for NNW, NNE and SW, respectively (Fig. 3). We analyzed the effects of velocity and direction of wind, the number of livestocks in each direction, and configuration of the ground on these results. The direction of wind was not regular, but the main direction of wind was toward east-northeast (ENE), and the average wind velocity was 1.6 m per second. It was difficult to estimate the effect of wind on direction of flight (Table 6). The number of livestock had some effects on the direction of flight, in direction of NNW, total 2,954 livestock had been living when this experiment was carried out. That number of livestock was much higher than those in other two directions. It was therefore highly likely that the mosquitoes flew to find a blood source (Fig. 4).
DISCUSSION
An. sinensis is well known as main vector of malaria transmission in Korea. This species comprises more than 80% of total mosquito population during the malaria transmission season. Other 6 species of Anopheline (An. lesteri, An. pullus. An. koreicus, An. sineroides, An. yatsushiroensis and An. lindesayi) also make up the population, but their proportion of these species is very low. In Korea, vivax malaria has been prevalent for many years, however, as the result of National Malaria Eradication Programme in conjunction with assistance from World Health Organization (WHO), the incidence of vivax malaria has rapidly decreased in Korea (National Malaria Eradication Service, 1966; Paik et al., 1988), and had been believed to be eradicated from Korea since the late 1970s. But more than 13,000 vivax malaria patients were detected since 2000 near demilitarized zone (DMZ). Most scientists suspected the North Korea as being the origin.
To clarify the origin, first we should elucidate the characteristics of Anopheline mosquitoes activity, because we have the buffering region called DMZ between North and South Korea, which are separated by 4 km. Therefore in order to study the flight range of female An. sinensis, we captured, marked them by orange and yellow fluorescent dyes, and released.
We recaptured 194 marked female mosquitoes, showing 1.52% of the recapture rate. There have been some differences of the recapture rates depending on the aim of dispersal experiment, the species of mosquitoes and configuration of dispersal area. Reisen et al. (1980) reported that the recapture rate of female An. culicifacies was 8.0%, and that of male was 5.9%, captured mainly in houses and cattle houses, therefore their recapture rate was somewhat higher than ours. On the other hand, Jaal & MacDonald (1992) reported that only 3 out of 8 species of Anopheline mosquitoes species were recaptured with the recapture rates of 3.42% for An. lesteri paraliae, 1.19% for An. subpictus, and 0.97% for An. vagus, in agreement with our results. In an another dispersal experiment by Kramer et al. (1995), Aedes dorsalis were released and recaptured during July and October, and 0.1% and 1.2% recapture rate was achieved in July and October, respectively. These results are very similar to ours. Kligler (1932) reported that An. sacharovi flew over 13 km from their larval habitat, and An. freeborni flies 42 km from release site to find a place for passing the winter, which has been the most longest ever reported (Freeborn, 1932). Ree et al. (1975) reported that Culex tritaeniorhynchus, which is the main vector of Japanese encephalitis in Korea, could fly 7.5 km in a night. Considering the mosquitoes captured in light trap which was set up at 7.5 km from the release site, C. tritaeniorhynchus females appeared to have the ability to fly much longer distance than 7.5 km. Bailey et al. (1965) reported that C. tarsalis can fly more than 28 km from the release site to find the place to pass winter, but most of them were found to be within 8.0 km from the release site. Kramer et al. (1995) described that most of Ae. dosalis were recaptured within 2.4 to 8.0 km from the release site. In our study, one An. sinensis was recaptured 20 km from the release site, suggesting that An. sinensis females have the ability to fly much longer distance with wind than 20 km.
Generally, movement of mosquito from one habitat to another is classified as being either active dispersal or passive dispersal. Active dispersal includes finding a resting place after bloodsucking, finding and mating place for fertilization, finding food such as nectar in the case of male mosquito, finding blood sources such as human or animals, and finding place for laying eggs in the case of female mosquito. On the other hand, passive dispersal is the movement of mosquito by ship, plane, train or wind. In the case of active dispersal, distances are different according to the species of mosquitoes. An quadrimaculatus in America and An. melas in West Africa can disperse only few hundred meters in closed areas. Eyles et al. (1945) reported that An. quadrimaculatus dispersed within 3 km, whereas Giglioli (1964, 1965) reported that An. melas moved regularly to find places to lay eggs and blood source within 1.6-4 km. In our study, 85% of the mosquitoes were recaptured within 6 km from the release point, therefore, it is likely An. sinensis can fly actively to find a new environment. Additionally, wind and geography also may have an influence. Total 58.8% (114/194) of marked An. sinensis moved to NNW, where plains and highways are well developed compared to the other two areas where dispersed.
Also, the number of livestock such as cows and pigs are higher in this area, and they influenced the movement of mosquitoes. It seemed that An. sinensis moved mainly to find a blood source. Kramer et al. (1995) reported that movement of Ae. dorsalis was influenced mainly by wind, but they didn't consider blood source and geographical diversity.
Since 170 female mosquitoes (87.6%) out of 194 marked mosquitoes were captured within 6 km from the release point, this flight radius represents the main activity area. An. sinensis was found to be able to fly at least 12 km during one night.