Dubey: Review of Neospora caninum and neosporosis in animals
Abstract
Neospora caninum is a coccidian parasite of animals. It is a major pathogen for cattle and dogs and it occasionally causes clinical infections in horses, goats, sheep, and deer. Domestic dogs are the only known definitive hosts for N. caninum. It is one of the most efficiently transmitted parasite of cattle and up to 90% of cattle in some herds are infected. Transplacental transmission is considered the major route of transmission of N. caninum in cattle. Neospora caninum is a major cause of abortion in cattle in many countries. To elicit protective immunity against abortion in cows that already harbor a latent infection is a major problem. This paper reviews information on biology, diagnosis, epidemiology and control of neosporosis in animals.
Key words: Neospora caninum, neosporosis, animals, cattle, dogs
INTRODUCTION
Since its first recognition in 1984 in dogs in Norway ( Bjerkås et al., 1984) and the description of a new genus and species Neospora caninum ( Dubey et al., 1988a), neosporosis has emerged as a serious disease of cattle and dogs worldwide. Additionally, clinical neosporosis has been reported in sheep, goats, deer, a rhinocerus, and horses and antibodies to N. caninum have been found in the sera of water buffaloes, red and gray foxes, coyotes, and camels, and felids.
BIOLOGY
Life Cycle
Dogs are both the intermediate and definitive host for N. caninum ( McAllister et al., 1998; Lindsay et al., 1999a, 1990b, 2001a; Basso et al., 2001a; Dubey et al., 2002). The life cycle is typified by 3 infectious stages: tachyzoites, tissue cysts, and oocysts ( Fig. 1 and Fig. 2). Tachyzoites and tissue cysts are the stages found in the intermediate hosts and they occur intracellularly ( Dubey et al., 2002). Tachyzoites are approximately 6 × 2 μm ( Fig. 1). Tissue cysts are often round or oval in shape, up to 107 μm long and are found primarily in the central nervous system (CNS). The tissue cyst wall is up to 4 μm thick and the enclosed bradyzoites are 7-8 × 2 μm. Thin-walled (0.3-1.0 μm) tissue cysts have been recently reported in muscles of cattle and dogs naturally-infected with a N. caninum-like parasite ( Peters et al., 2001a). These intramuscular tissue cysts have not been yet found in experimentally-infected animals ( Dubey et al., 2002). Domestic dogs are the only known definitive host for N. caninum. N. caninum unsporulated oocysts from experimentally-infected dogs were 11.7 × 11.3 (10.6-12.4 × 10.6-12.0) μm in size ( Lindsay et al., 1999a). N. caninum oocysts sporulate outside the host. N. caninum oocysts are morphologically similar to Toxoplasma gondii and Hammondia hammondi oocysts in cat feces and broadly resemble oocysts of Hammondia heydorni-like parasite in dog feces ( Dubey et al., 2002). At the present, nothing is known regarding the frequency of shedding of oocysts, the survival of the oocysts in the environment, and whether other canids are also definitive hosts for N. caninum.
Transmission
It is epidemiologically important to be able to identify N. caninum oocysts in dog feces. Microscopic examination alone will not be enough to identify N. caninum oocysts in dog feces. Methods have also been developed to genetically distinguish N. caninum oocysts from H. heydorni oocysts ( Hill et al., 2001; Šlapeta et al., 2002). Neospora caninum oocysts have been identified feces of only two naturally-infected dogs ( Basso et al., 2001a; Šlapeta et al., 2002).
Animal models
There are no suitable animal models at the present to perform bioassay to detect N. caninum oocysts in dog feces. Although interferon-gamma gene knockout (KO) mice are highly susceptible to parenteral inoculation with N. caninum tachyzoites and tissue cysts ( Dubey and Lindsay, 1996; Dubey et al., 1998a), they are less susceptible to parenteral or oral inoculation with oocysts. Gerbils ( Meriones unguiculatus) were susceptible to N. caninum infection with oocysts ( Dubey and Lindsay, 2000; Basso et al., 2001a; Schares et al., 2001). Another species of gerbils, Meriones tristrami and sand rats ( Psammoomys ubesus) also susceptible to tachyzoites infection ( Pipano et al., 2002).
Comparison between neosporosis and toxoplasmosis
A large body of knowledge has been generated concerning biology of N. caninum because of previous experience with the related parasite, Toxoplasma gondii. Although N. caninum and T. gondii are very closely related parasites structurally, genetically, and immunologically, caution should be used in making generalizations about N. caninum based on the biology of T. gondii because neosporosis and toxoplasmosis are biologically distinct diseases. Toxoplasma gondii is a major disease of sheep and humans and not of cattle, whereas neosporosis is a major disease in cattle, not of sheep and there is no evidence for human infection.
NEOSPOROSIS IN CATTLE
Bovine neosporosis has been reviewed in several papers ( Dubey and Lindsay, 1996; Wouda, 1998; Dubey, 1999; Anderson et al., 2000; Buxton et al., 2002; Dijkstra, 2002; Innes et al., 2002; Jenkins et al., 2002; Dubey, 2003). Therefore, most references on bovine neosporosis were omitted from this review.
Clinical signs
Neospora caninum causes abortion both dairy and beef cattle. Cows of any age may abort from 3 month gestation to term. Most neosporosis-induced abortions occur at 5-6 month gestation. Fetuses may die in utero, be resorbed, mummified, autolyzed, stillborn, born alive with clinical signs, or born clinically normal but chronically infected. Neosporosis-induced abortions occur year-round. Cows with N. caninum antibodies (seropositive) are more likely to abort than seronegative cows and this applies to both dairy and beef cattle. However, up to 95% of calves born congenitally-infected from seropositive dams remain clinically normal. The age of dam, lactation number, and history of abortion generally do not affect rate of congenital infection but there are reports indicating that in persistently infected cattle vertical transmission is more efficient in younger than older cows.
Clinical signs have only been reported in cattle younger than 2 month of age. Neospora caninum-infected calves may have neurologic signs, be underweight, unable to rise, or be born without clinical signs of disease. Hind limbs or forelimbs or both may be flexed or hyperextended. Neurologic examination may reveal ataxia, decreased patellar reflexes, and loss of conscious proprioception. Calves may have exophthalmia or asymmetrical appearance in the eyes. Neospora caninum occasionally causes birth defects including hydrocephalus and narrowing of the spinal cord.
Abortions may be epidemic or endemic ( Wouda et al., 1999a). In infected areas, as many as 33% of dairy cow fetuses have been reported to abort within a few months. Abortions were considered epidemic if more than 10% of cows at risk aborted within 6-8 weeks. A small proportion (< 5%) of cows have been reported to have repeated abortion due to neosporosis ( Anderson et al., 1995).
Prevalence
Neospora caninum infections have been reported from most parts of the world including Australia, New Zealand, Europe, Korea, Japan, Thailand, and the Americas. Neosporosis-associated bovine abortion and neonatal mortality has been reported from Argentina, Australia, Belgium, Brazil, Canada, Costa Rica, Denmark, France, Germany, Hungary, Ireland, Israel, Italy, Japan, Korea, Mexico, the Netherlands, New Zealand, Poland, Portugal, Spain, South Africa, Sweden, United Kingdom, USA, and Zimbabwe. Quantitative studies in the USA, New Zealand, the Netherlands, and Germany indicate that 12 to 42% of aborted fetuses from dairy cattle are infected with N. caninum ( Table 1).
Serologic prevalence in cattle varies, depending on the country, region, type of serologic test used, and cut-off level used to determine the exposure. In some dairies up to 87% of cows are seropositive.
Diagnosis
Examination of the serum from an aborting cow is only indicative of exposure to N. caninum and histologic examination of the fetus is necessary for a definitive diagnosis of neosporosis. The brain, heart, liver, placenta, and body fluids or blood serum are the best specimens for diagnosis and diagnostic rates are higher if multiple tissues are examined. Although lesions of neosporosis are found in several organs, fetal brain is the most consistently affected organ. Because most aborted fetuses are likely to be autolyzed, even semi-liquid brain tissue should be fixed in 10% buffered neutral formalin for histologic examination of hematoxylin and eosin (HE) stained sections. Immunohistochemistry is necessary because there are generally only a few N. caninum present in autolyzed tissues and these are often not visible in H and E stained sections. The most characteristic lesion of neosporosis is focal encephalitis characterized by necrosis and nonsuppurative inflammation ( Barr et al., 1991). Hepatitis is more common in epizootic than sporadic abortions. Lesions are also present in placenta but protozoa are difficult to find. The efficiency of the diagnosis by PCR is dependent on the laboratory, stage of the autolysis of the fetus, and sampling procedures. Although immunohistochemical demonstration of N. caninum in lesions is the best evidence for etiology of abortion at the present time, it is very insensitive. Neospora caninum DNA can be detected by PCR in formalin-fixed, paraffin-embedded bovine aborted brain tissue.
Several serologic tests can be used to detect N. caninum antibodies including various ELISAs, the indirect fluorescent antibody test (IFAT), and the Neospora agglutination test (NAT). Immunoblots are useful in detecting N. caninum-specific antibodies. Avidity-ELISAs designed to distinguish recent and chronic infections in cattle appear promising to distinguish endemic and epidemic abortion.
Finding N. caninum antibody in serum from the fetus can establish N. caninum infection, but a negative result is not informative because antibody synthesis in the fetus is dependent on the stage of gestation, level of exposure, and the time between infection and abortion. Even a low IFAT titer of 1:25 should be regarded as specific for N. caninum infection, especially in fetuses. Immunoblotting using N. caninum specific antigen improves diagnosis. Although blood serum or any body fluid from the fetus may be used for serologic diagnosis, peritoneal fluid is better than other body fluids. In calves, presuckling serum can be submitted for diagnosis of congenital infection.
The definitive antibody level that should be considered diagnostic for neosporosis has not been established for bovines because of the uncertainty of serologic diagnosis in chronically infected animals and the availability of sera from noninfected cattle. In serological assays, titer and absorbance values are dependant on antigen composition, secondary antibodies and other reagents. Further, cut-off levels can be arbitrarily selected to provide sensitivity and specificity requested for a particular application. The age and class of an animal may also affect selection of a cut-off level. Although N. caninum is closely related to Toxoplasma gondii, Sarcocystis species, and other apicomplexans, cross reactivity has not been a major issue. Antibody titers in general are higher in cattle that have aborted due to neosporosis than those with normal pregnancy; however, titers in individual cows cannot determine etiology of abortions.
Toxoplasma gondii and Sarcocystis cruzi are 2 other protozoans that should be considered in the differential diagnosis of protozoal abortion in cattle. Immunohistochemical and detection of parasite DNA by PCR can distinguish them from N. caninum. Sarcocystis cruzi forms schizonts in vascular endothelium and is rarely (< 0.1%) found in aborted fetal brains, whereas N. caninum is usually located in extravascular tissues. Additionally, there are no immature schizonts in N. caninum infection in contrast with S. cruzi infections. Infection by T. gondii in bovine fetuses is rare ( Canada et al., 2002b).
Attempts at isolation of viable N. caninum ( Table 2) by bioassay in mice or cell culture have been largely unsuccessful and little is known of the antigenic variability among isolates of N. caninum, especially isolates from healthy animals. Many attempts to isolate viable N. caninum were unsuccessful because most N. caninum in fetuses die with the host. It is easier to isolate N. caninum from neural tissues of congenitally-infected full term calves ( Table 2) because tissue cysts are likely to be present and tissue cysts are relatively more resistant to autolysis than tachyzoites.
Pathogenesis of abortion
Cows with N. caninum antibodies (seropositive) are more likely to abort than seronegative cows. There is a rise in antibody titers 4 to 5 months before parturition. These observations strongly suggest reactivation of latent infection. Little is known of the mechanism of reactivation. It is likely that there is parasitemia during pregnancy leading to fetal infection. However, N. caninum has never been identified in histologic sections of adult cows and there is a single report of isolation of viable N. caninum from the brain of an adult cow ( Sawada et al., 2000). Although it is reasonable to speculate that pregnancy-induced immune-suppression may reactivate latent tissue cysts of N. caninum, such a mechanism has not been demonstrated for neosporosis. Little is known at the present time concerning the pathogenesis of infection in cows following infection with a natural route (oral) and using the naturally transmitted stage of the parasite (oocyst) although oocysts are infective to cattle by the oral route. Results of experiments with dairy or beef cows inoculated parenterally with N. caninum tachyzoites indicate that the fetus can become infected and sometimes diseased by 4 weeks after inoculation of the parasite. Even at the early stage, there were lesions in placenta and the CNS and encephalitis was the predominent lesion. The extent of damage to placenta directly due to multiplication of N. caninum and by immune response is not known ( Buxton et al., 2002). Gestational age may determine the outcome of infection. Fetuses infected early in pregnancy are likely to die.
Neospora caninum is a primary pathogen in cattle
Neospora caninum is one of the most efficiently transplacentally-transmitted organisms in cattle. In some herds up to 90% of cattle are infected, and most calves born congenitally-infected with N. caninum remain healthy. Therefore, there is a debate concerning whether N. caninum causes abortion in cattle or whether it is a bystander ( Thurmond et al., 1999). All evidence at the present time indicates that N. caninum is a primary pathogen. However, there is a need to distinguish clinical and nonclinical N. caninum infections. In the initial reports by Anderson et al. ( 1991) and Barr et al. ( 1991), the following criteria were established to diagnose N. caninum abortion. First, the fetuses had lesions, mostly encephalitis. Second, N. caninum was found in lesions. In my opinion, identification of organisms in lesions is a good evidence for the etiology of the lesions. Additionally, abortion has been induced in experimentally-infected cattle. The percentage of fetuses that have histologically detectable N. caninum and no lesions is unknown. Therefore, detection of N. caninum DNA alone in aborted fetuses may not be enough to establish cause and effect relationship. Host factors (cytokines, hormones) that may contribute to pathogenesis of N. caninum infection have not yet been established ( Buxton et al., 2002; Innes et al., 2002).
Transmission
There is no cow to cow transmission of N. caninum. In the study by Anderson et al. ( 1997), 25 seronegative heifers were housed with 25 seropositive heifers since birth and their progeny were evaluated for N. caninum infection. The seronegative heifers remained seronegative and gave birth to calves not infected with N. caninum. Seropositive heifers remained clinically normal but gave birth to congenitally-infected calves. Seven of these congenitally-infected calves were necropsied; all had histologic evidence of N. caninum infection and 4 were recumbent. Although most N. caninum infections in cattle are transmitted transplacentally, postnatal rates have been variable depending on the region of the country, type of test used and cut-off values used. It is unlikely that N. caninum is transmitted venereally or by embryo transfer in cattle and embryo transfer is even recommended as a method of control to prevent vertical transmission ( Baillargeon et al., 2001). These researchers conducted an important study on embryo transfer and N. caninum infection involving 87 recipient cows or heifers. Neospora caninum infection was not demonstrable in any of the 70 fetuses or calves born to seronegative cows transplanted embryos from seropositive donors whereas 5 of 6 calves resulting from embryo transfer from seronegative donors to seropositive recipients were infected with N. caninum. Landmann et al. ( 2002) confirmed these findings and showed that commercially used embryo transfer procedures also prevented transfer of N. caninum from seropositive cows to seronegative recipients. Additionally, preimplanted bovine embryos are resistant to N. caninum invasion ( Bielanski et al., 2002). Lactogenic transmission of N. caninum has been demonstrated experimentally in newborn calves fed colostrum spiked with tachyzoites, but there is no evidence that it occurs naturally ( Davison et al., 2001); dogs fed milk spiked with N. caninum tachyzoites did not shed oocysts ( Dijkstra et al., 2001).
Economic impact
There are no firm data on the economic losses due to neosporosis to cattle industry anywhere in the world but losses are estimated in millions of dollars. As many as 42% of cows may abort due to neosporosis, the economic impact will depend on the direct cost and value of fetuses lost. Indirect costs include professional help and costs associated with establishing diagnosis, rebreeding, possible loss of milk yield, and replacement costs if aborted cows are culled ( Thurmond and Hietala, 1996, 1997; Hernandez et al., 2001). In general, less is known of the causes of abortion in beef cattle than in dairy cattle because of the difficulty of finding small fetuses expelled in the first trimester. Therefore, there are no accurate assessments of Neospora-induced losses in beef cattle. Because clinical disease has not been reported in calves older than 2 months of age, there is no direct evidence of N. caninum-associated morbidity in adult cattle. However, in a sero-epidemiological study, Barling et al. ( 2000) found a positive association between N. caninum antibody status of the calf and weight gain and projected a $15.62 loss per calf.
Epidemiology and control
As stated earlier, N. caninum is efficiently transmitted vertically in cattle, perhaps for several generations. Therefore, culling is the only way at present to prevent this transmission from cow to heifer. Until the discovery of the resistant stage (oocyst) of N. caninum, horizontal transmission was unexplained. Evidence is accumulating that transmission by oocysts may be more prevalent than initially realized, at least in some parts of the world. Storms of abortions, abortion in recently infected cattle as evidenced by low avidity antibodies, and seroconversion in a high portion of cattle, indicate transmission of N. caninum by fecal-oral route. Postnatal transmission also occurs in herds not experiencing storms of abortions and seroepidemiological data support the role of the dog in the life cycle of N. caninum.
How dogs become infected in nature is not known. Small animals that dogs might prey or consume naturally have not been identified as a natural host for N. caninum. Consumption of aborted bovine fetuses does not appear to be an important source of N. caninum infection in dogs. The consumption of placental membranes may be a source of N. caninum infection in dogs because the parasite has been found in naturally-infected placentas and dogs fed placentas shed N. caninum oocysts. Little is known at present regarding the frequency of shedding of N. caninum oocysts by canids in nature, the resistance of the oocysts, and whether dogs shed oocysts more than once. Until more definitive hosts of N. caninum are found, dogs should not be allowed to eat aborted fetuses, fetal membranes or dead calves. Drugs that will prevent transmission of the parasite for the dam to the fetus are unknown but research is continuing in this area.
There is no proven vaccine to prevent N. caninum abortion in cattle ( Innes et al., 2002). However, encouraging results have been obtained in mice vaccinated with killed parasites or their products. Vaccination of mice before pregnancy with killed N. caninum tachyzoites blocked transplacental transfer of N. caninum when mice were challenged during pregnancy demonstrated protective immunity to congenital transfer of N. caninum infection has been demonstrated in experimentally-infected cattle ( Innes et al., 2001b). As mentioned earlier, protective immunity appears to develop which reduces or prevents subsequent abortion after a primary infection and resultant abortion in cattle ( Anderson et al., 1995; McAllister et al., 2000). This protective immunity appears to be more effective in cows that are infected again from an exogenous source (oocysts) than in cows that relapse from an endogenous infection ( Trees and Williams, 2003). Therefore, inducing protective immunity in cows that are already infected naturally is a problem.
NEOSPOROSIS IN DOGS
Neosporosis was first recognized in dogs in Norway by Bjerkås et al. ( 1984). Retrospectively, the earliest record of neosporosis in dogs was an outbreak in 1957 in the U.S. ( Dubey et al., 1990a). Worldwide reports of clinical and subclinical infections were summarized by Dubey and Lindsay ( 1996) and Lindsay and Dubey ( 2000) and these references are not repeated here. Since then, antibodies to N. caninum were reported in 121 of 320 (37.8%) dogs from Argentina ( Basso et al., 2001b), 22% of 200 dogs from New Zealand ( Reichel et al., 1998), 10% of 150 dogs from Turkey ( Coškun et al., 2000), 6.7% of 163 dogs from Brazil ( Mineo et al., 2001), 10% of 500 pet dogs and 25% of 611 stray dogs from Brazil ( Gennari et al., 2002), 6.4% of 1,058 dogs from Italy ( Cringoli et al., 2002), and in 12.% of 120 urban and 26% of 81 rural dogs from Chile ( Patitucci et al., 2001). Klein and Müller ( 2001) reported N. caninum antibodies in 4% of 50 dogs in Germany without clinical signs and 13% of 200 dogs with clinical signs. Antibodies to N. caninum were found in 21.6% of 134 dogs from cattle farms in Parana, Brazil ( de Souza et al., 2002). Three surveys compared the prevalence of N. caninum in urban and rural dogs. Sawada et al. ( 1998) reported N. caninum antibodies in 31% of 48 dogs from dairy farms and 7% of 198 dogs from urban areas in Japan. Wouda et al. ( 1999b) reported a higher prevalence in farm dogs (23.6% of 152) versus urban dogs (5.5% of 344) from the Netherlands. Basso et al. ( 2001b) reported higher seroprevalence in dogs from dairy farms (48% of 125) and beef farms (54.2% of 35) than in dogs from urban areas (22.2% of 160) of Argentina. In addition to clinical infections summarized by Dubey and Lindsay ( 1996) and Lindsay and Dubey ( 2000), Boydell and Brogan ( 2000), Cantile and Arispici ( 2002) reported clinical neurologic neosporosis in dogs. Four cases of cutaneous neosporosis were recently reported, including mixed infection with Leishmania sp. in 1 dog ( Tarantino et al., 2001; Perlé et al., 2001; Ordeix et al., 2002). Barber and Trees ( 1998) reported an interesting seroepidemiologic study of N. caninum infections in 373 breeding dogs in U.K. Fifty (13.4%) of 373 bitches had an IFAT titer of > 1:50. Initially, approximately 50% pups born were seropositive and approximately 25% of pups born from seropositive dams developed neosporosis-like illness. Three bitches produced successive litters of N. caninum infected pups. In subsequent breeding only 3 of 118 pups from seronegative dams were seropositive. Thus, vertical transmission alone could not maintain the parasite in dogs ( Barber and Trees, 1998). The most severe cases of neosporosis occur in young, congenitally infected pups. Young dogs develop hind limb paresis that develops into a progressive paralysis. Neurologic signs are dependent on the site parasitized. The hind limbs are more severely affected than the front limbs and often in rigid hyperextention. Other dysfunctions which occur include difficulty in swallowing, paralysis of the jaw, muscle flaccidity, muscle atrophy and even heart failure. Dogs with hind limb paralysis may be alert and survive for months.
The disease may be localized or generalized and virtually all organs may be involved including the skin. Dermatitis may be severe involving enormous numbers of N. caninum. Dogs of any age can be affected. Fatal neosporosis has been reported in eight to 15-year-old dogs.
Subclinically infected bitches can transmit the parasite to their fetuses, and successive litters from the same bitch may be born infected. Whether there is breed predisposition and differential sex susceptibility to neosporosis in dogs is not known. Most described cases have been in Labrador retrievers, Boxers, Greyhounds, Golden retrievers, and Basset hounds.
Neospora caninum has been isolated several times from dogs, mostly from dogs with neuromuscular signs ( Table 3). Nothing is known of the isolates of N. caninum from dogs without any clinical signs.
NEOSPOROSIS IN WATER BUFFALOES
Water buffaloes ( Bubalus bubalis) are important to the economy of several countries including Brazil, India, Italy, and Vietnam. Prevalence of antibodies indicates that buffaloes in four countries were exposed to N. caninum ( Table 4). Little is known of the role of N. caninum in causing abortion in buffaloes. Guarino et al. ( 2000) reported lesions and Neospora-like tissue cysts in 2 of 4 fetuses aborted from 2 dairy farms in Italy; this report needs confirmation.
NEOSPOROSIS IN SHEEP
Neospora caninum was first diagnosed in a congenitally-infected lamb in England, ( Dubey et al., 1990b). Historically, this was the first record of N. caninum-like infection in a ruminant ( Hartley and Bridge, 1975). The lamb was born weak, partially ataxic, and died at one week of age. There was unilateral reduction of gray matter in the ventral horn and local cavitation. This case was published because congenital deformities due to toxoplasmosis had not been seen in any animal (other than humans), only tissue cysts and not tachyzoites were seen. I had known of this case even before it was published. After the discovery of the N. caninum in dogs in 1988, I contacted William Hartley and retrospectively examined tissues of this lamb and an identical case in a calf that was added as an addendum to the case in lamb ( Hartley and Bridge, 1975); the calf was from Australia. Subsequently, clinical ovine neosporosis was reported from Japan. Kobayashi et al. ( 2001) found naturally-infected neosporosis in ewe and her twin fetus in Japan. Focal encephalitis and thick-walled N. caninum tissue cysts were seen in all three animals. Koyama et al. ( 2001) obtained the first isolate of N. caninum from another adult ewe.
Little is known of the seroprevalence of N. caninum in naturally exposed sheep and evidence for N. caninum as an ovine abortifacient in the field has not been found ( Otter et al., 1997). In a recent survey study from U.K., only 3 of 660 ewes that had aborted had antibodies to N. caninum ( Helmick et al., 2002). Pregnant sheep, however, are highly susceptible to experimental infection with N. caninum tachyzoites ( Dubey and Lindsay, 1990; McAllister et al., 1996; Buxton et al., 1997b, 1998; Jolley et al., 1999; Innes et al., 2001a) and are an alternative ruminant model for bovine neosporosis. Sheep can also be infected orally with N. caninum oocysts ( O'Handley et al., 2002).
NEOSPOROSIS IN GOATS
Abortion and neonatal mortality associated with N. caninum was reported in pygmy goats in the U.S. ( Barr et al., 1992; Dubey et al., 1992), in a dairy goat herd in Costa Rica ( Dubey et al., 1996a), and in a 3-day-old dairy goat from Brazil ( Corbellini et al., 2001). Antibodies to N. caninum were found in 5 of 77 dairy goats in Costa Rica that had aborted. Little is known of the seroprevalence of N. caninum antibodies in goats. Ooi et al. ( 2000) did not find N. caninum antibodies in 24 goats from Taiwan. Pygmy goats, however, are quite susceptible to experimental N. caninum infection. Goats inoculated N. caninum during pregnancy aborted N. caninum-infected fetuses ( Lindsay et al., 1995).
NEOSPOROSIS IN HORSES
Neospora-like parasites have been found in tissues of two aborted foals ( Dubey and Porterfield, 1990; Pronost et al., 1999), a congenitally-infected foal ( Lindsay et al., 1996b) and five adult horses ( Gray et al., 1996; Daft et al., 1996; Marsh et al., 1996; Hamir et al., 1998; Cheadle et al., 1999b). Marsh et al. ( 1998) proposed a new name, Neospora hughesi, for the parasite in the horse they reported in 1996. Three isolates of N. hughesi from adult horses have been reported ( Marsh et al., 1998; Cheadle et al., 1999b; Dubey et al., 2001). Molecular and biological characteristics of these three isolates have been reported ( Cheadle et al., 1999a; Walsh et al., 2000; Dubey et al., 2001; Marsh et al., 2001).
Tissue cysts of N. hughesi were smaller than N. caninum with thinner cyst walls (less than 1.0 µm thick), and bradyzoites were smaller than those of N. caninum ( Marsh et al., 1998). It is, however, not clear at the present time whether N. hughesi is the sole species of Neospora that infects horses. Thick walled tissue cysts, characteristics of N. caninum were reported from a horse from California ( Daft et al., 1996) and a congenitally infected foal from Wisconsin ( Lindsay et al., 1996b). Seroprevalence of Neospora infections in horses is summarized in Table 5. In two surveys or horses in France and the U.S. the seroprevalence by NAT was 23%.
NEOSPORA INFECTIONS IN HUMANS
Because two rhesus monkeys ( Macaca mulata) have been successfully infected with N. caninum ( Barr et al., 1994), there is a concern about the zoonotic potential of N. caninum. However, at present there is no evidence that N. caninum successfully infects humans. Graham et al. ( 1999) did not find N. caninum antibodies in 1:160 dilution of sera from 199 blood donors and 48 agricultural workers in Northern Ireland. Petersen et al. ( 1999) did not find antibodies to N. caninum in 76 women with history of abortion using ELISA, immunoblotting, and IFAT. Low level IFAT (1:100) antibodies were reported by Tranas et al. ( 1999) in 69 (6.7%) of 1,029 sera from blood donors in California, however, all sera were negative at 1:200 dilution. Nam et al. ( 1998) reported antibodies to N. caninum in human sera using the immunoblots; however, these sera were negative by NAT. Trees and Williams. ( 2000) found low (IFAT < 1:200) in 2 of 500 (400 farm workers and 100 women with recurrent abortions) people in England. In conclusion, there is no evidence that N. caninum infection is zoonotic.
NEOSPOROSIS IN WILDLIFE
Neosporosis was diagnosed at necropsy in two black-tailed deer ( Odocoileus hemionus columbianus) found dead in the wild in California ( Woods et al., 1994), in a captive deer in zoos in France ( Dubey et al., 1996b, and Germany ( Peters et al., 2001b. Neospora caninum tissue cysts were found in the brain of a full term stillborn deer ( Cervus eldi siamensis) in a Paris Zoo ( Dubey et al., 1996b. Peters et al. ( 2001b) reported antibodies to N. caninum in fetal fluid and N. caninum DNA by PCR in the brain, heart, lung, liver and spleen of 2 full term twin calves of antelope ( Tragelaphus imberberis) in a zoo in Hannover, Germany. Neosporosis was reported in a rhinoceros ( Ceratotherium simum) calf ( Williams et al., 2002). The animal died suddenly, considered to be due to myocarditis. Rhinoceroses are related to equids, all belonging to the order Perissodactyla. Finding of antibodies in 40% of wild deer (Table 9) indicates there may be a sylvatic cycle of N. caninum. A large percentage of these deer had high titered (> 1:1,600) N. caninum antibodies and the prevalence did not increase with age indicating that congenital transmission ( Dubey et al., 1999a). Similarly, red foxes in Belgium had a high seroprevalence of N. caninum. Schares et al. ( 2001) presented evidence of congenital infection of N. caninum in foxes.
REFERENCES
1. Alméria S, Ferrer D, Pabón M, Castell J, Maas S. Red foxes ( Vulpes vulpes) are a natural intermediate host of Neospora caninum. Vet Parasitol 2002;107:287-294. PMID: 12163240.  2. Anderson ML, Blanchard PC, Barr BC, Dubey JP, Hoffmann RL, Conrad PA. Neospora-like protozoan infection as a major cause of abortion in California dairy cattle. J Am Vet Med Assoc 1991;198:241-244. PMID: 2004983. 3. Anderson ML, Palmer CW, Thurmond MC, et al. Evaluation of abortions in cattle attributable to neosporosis in selected dairy herds in California. J Am Vet Med Assoc 1995;207:1206-1210. PMID: 7559072. 4. Anderson ML, Reynolds JP, Rowe JD, et al. Evidence of vertical transmission of Neospora sp infection in dairy cattle. J Am Vet Med Assoc 1997;210:1169-1172. PMID: 9108925. 5. Anderson ML, Andrianarivo AG, Conrad PA. Neosporosis in cattle. Anim Reprod Sci 2000;60-61:417-431. PMID: 10844212. 6. Baillargeon P, Fecteau G, Pare J, Lamothe P, Sauve R. Evaluation of the embryo transfer procedure proposed by the International Embryo Transfer Society as a method of controlling vertical transmission of Neospora caninum in cattle. J Am Vet Med Assoc 2001;218:1803-1806. PMID: 11394835. 7. Barber JS, Holmdahl OJM, Owen MR, Guy F, Uggla A, Trees AJ. Characterization of the first European isolate of Neospora caninum (Dubey, Carpenter, Speer, Topper and Uggla). Parasitology 1995;111:563-568. PMID: 8559588. 8. Barber JS, Gasser RB, Ellis J, Reichel MP, McMillan D, Trees AJ. Prevalence of antibodies to Neospora caninum in different canid populations. J Parasitol 1997;83:1056-1058. PMID: 9406778. 9. Barber JS, Trees AJ. Naturally occurring vertical transmission of Neospora caninum in dogs. Int J Parasitol 1998;28:57-64. PMID: 9504335. 10. Barling KS, McNeill JW, Thompson JA, et al. Association of serologic status for Neospora caninum with postweaning weight gain and carcass measurements in beef calves. J Am Vet Med Assoc 2000;217:1356-1360. PMID: 11061390. 11. Barr BC, Anderson ML, Dubey JP, Conrad PA. Neospora-like protozoal infections associated with bovine abortions. Vet Pathol 1991;28:110-116. PMID: 2063512. 12. Barr BC, Anderson ML, Woods LW, Dubey JP, Conrad PA. Neospora-like protozoal infections associated with abortion in goats. J Vet Diagn Invest 1992;4:365-367. PMID: 1515507. 13. Barr BC, Conrad PA, Breitmeyer R, et al. Congenital Neospora infection in calves born from cows that had previously aborted Neospora-infected fetuses: four cases (1990-1992). J Am Vet Med Assoc 1993;202:113-117. PMID: 8420896. 14. Barr BC, Conrad PA, Sverlow KW, Tarantal AF, Hendrickx AG. Experimental fetal and transplacental Neospora infection in the nonhuman primate. Laboratory Investigation 1994;71:236-242. PMID: 8078303. 15. Basso W, Venturini L, Venturini MC, et al. First isolation of Neospora caninum from the feces of a naturally infected dog. J Parasitol 2001a;87:612-618. PMID: 11426726. 16. Basso W, Venturini L, Venturini MC, et al. Prevalence of Neospora caninum infection in dogs from beef-cattle farms, dairy farms, and from urban areas of Argentina. J Parasitol 2001b;87:906-907. PMID: 11534656. 17. Bielanski A, Robinson J, Phipps-Todd B. Effect of Neospora caninum on in vitro development of preimplantation stage bovine embryos and adherence to the zona pellucida. Vet Rec 2002;150:316-318. PMID: 11913589. 18. Bjerkås I, Mohn SF, Presthus J. Unidentified cyst-forming sporozoon causing encephalomyelitis and myositis in dogs. Z Parasitenk 1984;70:271-274. PMID: 6426185. 19. Boulton JG, Gill PA, Cook RW, Fraser GC, Harper PAW, Dubey JP. Bovine Neospora abortion in north-eastern New South Wales. Aust Vet J 1995;72:119-120. PMID: 7611986. 20. Boydell P, Brogan N. Horner's syndrome associated with Neospora infection. J Small Anim Pract 2000;41:571-572. PMID: 11138858. 21. Buxton D, Maley SW, Pastoret PP, Brochier B, Innes EA. Examination of red foxes ( Vulpes vulpes) from Belgium for antibody to Neospora caninum and Toxoplasma gondii. Vet Rec 1997a;141:308-309. PMID: 9330477. 22. Buxton D, Maley SW, Thomson KM, Trees AJ, Innes EA. Experimental infection of non-pregnant and pregnant sheep with Neospora caninum. J Comp Pathol 1997b;117:1-16. PMID: 9263840. 23. Buxton D, Maley SW, Wright S, Thomson KM, Rae AG, Innes EA. The pathogenesis of experimental neosporosis in pregnant sheep. J Comp Pathol 1998;118:267-279. PMID: 9651804. 24. Buxton D, McAllister M, Dubey JP. The comparative pathogenesis of neosporosis. Trends Parasitol 2002;18:546-552. PMID: 12482540. 25. Canada N, Meireles CS, Rocha A, et al. First Portuguese isolate of Neospora caninum from an aborted fetus from a dairy herd with endemic neosporosis. Vet Parasitol 2002a;110:11-15. PMID: 12446085. 26. Canada N, Meireles CS, Rocha A, Correia da Costa JM, Erickson MW, Dubey JP. Isolation of viable Toxoplasma gondii from naturally-infected aborted bovine fetuses. J Parasitol 2002b;88:1247-1248. PMID: 12537120. 27. Cantile C, Arispici M. Necrotizing cerebellitis due to Neospora caninum infection in an old dog. J Vet Med 2002;49:47-50. 28. Cheadle MA, Spencer JA, Blackburn BL. Seroprevalences of Neospora caninum and Toxoplasma gondii in nondomestic felids from southern Africa. J Zoo Wildlife Med 1999a;30:248-251.
29. Cheadle MA, Lindsay DS, Rowe S, et al. Prevalence of antibodies to Neospora caninum in horses from Alabama and characterisation of an isolate recovered from a naturally infected horse. Int J Parasitol 1999b;29:1537-1543. PMID: 10608440. 30. Conrad PA, Barr BC, Sverlow KW, et al. In vitro isolation and characterization of a Neospora sp. from aborted bovine foetuses. Parasitology 1993;106:239-249. PMID: 8488061. 31. Corbellini LG, Colodel EM, Driemeier D. Granulomatous encephalitis in a neurologically impaired goat kid associated with degeneration of Neospora caninum tissue cysts. J Vet Diagn Invest 2001;13:416-419. PMID: 11580064. 32. Corbellini LG, Driemeier D, Cruz CFE, Gondim LFP, Wald V. Neosporosis as a cause of abortion in dairy cattle in Rio Grande do Sul, southern Brazil. Vet Parasitol 2002;103:195-202. PMID: 11750112. 33. Cringoli G, Rinaldi E, Capuano F, Baldi L, Veneziano V, Capelli G. Serological survey of Neospora caninum and Leishmania infantum co-infection in dogs. Vet Parasitol 2002;106:307-313. PMID: 12079736. 34. Cuddon P, Lin DS, Bowman DD, et al. Neospora caninum infection in English springer spaniel littermates: Diagnostic evaluation and organism isolation. J Vet Intern Med 1992;6:325-332. PMID: 1484374. 35. Çoškun SZ, Aydyn L, Bauer C. Seroprevalence of Neospora caninum infection in domestic dogs in Turkey. Vet Rec 2000;146:649. PMID: 10872789. 36. Daft BM, Barr BC, Collins N, Sverlow K. Neospora encephalomyelitis and polyradiculoneuritis in an aged mare with Cushing's disease. Equine Vet J 1996;28:240-243.
37. Davison HC, Guy F, Trees AJ, et al. In vitro isolation of Neospora caninum from a stillborn calf in the UK. Res Vet Sci 1999;67:103-105. PMID: 10425249. 38. Davison HC, Guy CS, McGarry JW, Guy F, Williams DJL, Trees AJ. Experimental studies on the transmission of Neospora caninum between cattle. Res Vet Sci 2001;70:163-168. PMID: 11356096. 39. de Souza SLP, Guimares JS, Ferreira F, Dubey JP, Gennari SM. Prevalence of Neospora caninum antibodies in dogs from dairy cattle farms in Parana, Brazil. J Parasitol 2002;88:408-409. PMID: 12054023. 40. Dijkstra T, Eysker M, Schares G, Conraths FJ, Wouda W, Barkema HW. Dogs shed Neospora caninum oocysts after ingestion of naturally infected bovine placenta but not after ingestion of colostrum spiked with Neospora caninum tachyzoites. Int J Parasitol 2001;31:747-752. PMID: 11403764. 41. Dijkstra T. Horizontal and vertical transmission of Neospora caninum. 2002, the Netherlands. Univ. Utrecht. pp 1-140 PhD Thesis.
42. Dubey JP, Carpenter JL, Speer CA, Topper MJ, Uggla A. Newly recognized fatal protozoan disease of dogs. J Am Vet Med Assoc 1988a;192:1269-1285. PMID: 3391851. 43. Dubey JP, Hattel AL, Lindsay DS, Topper MJ. Neonatal Neospora caninum infection in dogs: Isolation of the causative agent and experimental transmission. J Am Vet MEd Assoc 1988b;193:1259-1263. PMID: 3144521. 44. Dubey JP, Lindsay DS. Neospora caninum induced abortion in sheep. J Vet Diagn Invest 1990;2:230-233. PMID: 2094452. 45. Dubey JP, Porterfield ML. Neospora caninum (Apicomplexa) in an aborted equine fetus. J Parasitol 1990;76:732-734. PMID: 2213418. 46. Dubey JP, Koestner A, Piper RC. Repeated transplacental transmission of Neospora caninum in dogs. J Am Vet Med Assoc 1990a;197:857-860. PMID: 2228766. 47. Dubey JP, Hartley WJ, Lindsay DS, Topper MJ. Fatal congenital Neospora caninum infection in a lamb. J Parasitol 1990b;76:127-130. PMID: 2299518. 48. Dubey JP, Acland HM, Hamir AN. Neospora caninum (Apicomplexa) in a stillborn goat. J Parasitol 1992;78:532-534. PMID: 1597802. 49. Dubey JP, Lindsay DS. A review of Neospora caninum and neosporosis. Vet Parasitol 1996;67:1-59. PMID: 9011014. 50. Dubey JP, Morales JA, Villalobos P, Lindsay DS, Blagburn BL, Topper MJ. Neosporosis-associated abortion in a dairy goat. J Am Vet Med Assoc 1996a;208:263-265. PMID: 8567387. 51. Dubey JP, Rigoulet J, Lagourette P, George C, Longeart L, LeNet JL. Fatal transplacental neosporosis in a deer ( Cervus eldi siamensis). J Parasitol 1996b;82:338-339. PMID: 8604111. 52. Dubey JP, Dorough KR, Jenkins MC, et al. Canine neosporosis: clinical signs, diagnosis, treatment and isolation of Neospora caninum in mice and cell culture. Int J Parasitol 1998a;28:1293-1304. PMID: 9762578. 53. Dubey JP, Romand S, Hilali M, Kwok OCH, Thulliez P. Seroprevalence of antibodies to Neospora caninum and Toxoplasma gondii in water buffaloes ( Bubalus bubalis) from Egypt. Int J Parasitol 1998b;28:527-529. PMID: 9559371. 54. Dubey JP, Hollis K, Romand S, et al. High prevalence of antibodies to Neospora caninum in white-tailed deer ( Odocoileus virginianus). Int J Parasitol 1999a;29:1709-1711. PMID: 10608458. 55. Dubey JP, Romand S, Thulliez P, Kwok OCH, Shen SK, Gamble HR. Prevalance of antibodies to Neospora caninum in horses in North America. J Parasitol 1999b;85:968-969. PMID: 10577738. 56. Dubey JP, Kerber CE, Granstrom DE. Serologic prevalence of Sarcocystis neurona, Toxoplasma gondii, and Neospora caninum in horses in Brazil. J Am Vet Med Assoc 1999c;215:970-972. PMID: 10511862. 57. Dubey JP, Venturini MC, Venturini L, McKinney J, Pecoraro M. Prevalence of antibodies to Sarcocystis neurona, Toxoplasma gondii, and Neospora caninum in horses from Argentina. Vet Parasitol 1999d;86:59-62. PMID: 10489203. 58. Dubey JP. Recent advances in Neospora and neosporosis. Vet Parasitol 1999;84:349-367. PMID: 10456423. 59. Dubey JP, Lindsay DS. Gerbils ( Meriones unguiculatus) are highly susceptible to oral infection with Neospora caninum oocyst. Parasitol Res 2000;86:165-168. PMID: 10685849. 60. Dubey JP, Liddell S, Mattson D, Speer CA, Howe DK, Jenkins MC. Characterization of the Oregon isolate of Neospora hughesi from a horse. J Parasitol 2001;87:345-353. PMID: 11318565. 61. Dubey JP, Barr BC, Barta JR, et al. Redescription of Neospora caninum and its differentiation from related coccidia. Int J Parasitol 2002;32:929-946. PMID: 12076623. 62. Dubey JP. Neosporosis in cattle. J Parasitol 2003;89(Supplement):in press. 63. Fioretti DP, Rosignoli L, Ricci G, Moretti A, Pasquali P, Polidori GA. Neospora caninum infection in a clinically healthy calf: parasitological study and serological follow-up. J Vet Med B 2000;47:47-53. 64. Fujii TU, Kasai N, Nishi SM, Dubey JP, Gennari SM. Seroprevalence of Neospora caninum in female water buffaloes ( Bubalus bubalis) from the southeastern region of Brazil. Vet Parasitol 2001;99:331-334. PMID: 11511420. 65. Gennari SM, Yai LEO, D'Auria SNR, et al. Occurrence of Neospora caninum antibodies in sera from dogs of the city of São Paulo, Brazil. Vet Parasitol 2002;106:177-179. PMID: 12031819. 66. Gondim LFP, Pinheiro AM, Santos POM, et al. Isolation of Neospora caninum from the brain of a naturally infected dog and production of encysted bradyzoites in gerbils. Vet Parasitol 2001;101:1-7. PMID: 11587828. 67. Gondim LFP, Gao L, McAllister MM. Improved production of Neospora caninum oocysts, cyclical oral transmission between dogs and cattle, and in vitro isolation from oocysts. J Parasitol 2002;88:1159-1163. PMID: 12537111. 68. Graham DA, Calvert V, Whyte M, Marks J. Absence of serological evidence for human Neospora caninum infection. Vet Rec 1999;144:672-673. PMID: 10404607. 69. Gray ML, Harmon BG, Sales L, Dubey JP. Visceral neosporosis in a 10-year-old horse. J Vet Diagn Invest 1996;8:130-133. PMID: 9026072. 70. Guarino A, Fusco G, Savini G, Di Francesco G, Cringoli G. Neosporosis in water buffalo ( Bubalus bubalis) in southern Italy. Vet Parasitol 2000;91:15-21. PMID: 10889356. 71. Gupta GD, Lakritz J, Kim JH, Kim DY, Kim JK, Marsh AE. Seroprevalence of Neospora, Toxoplasma gondii, and Sarcocystis neurona antibodies in horses from Jeju island, South Korea. Vet Parasitol 2002;106:193-201. PMID: 12062508. 72. Hamir AN, Tornquist SJ, Gerros TC, Topper MJ, Dubey JP. Neospora caninum - associated equine protozoal myeloencephalitis. Vet Parasitol 1998;79:269-274. PMID: 9831950. 73. Hartley WJ, Bridge PS. A case of suspected congenital Toxoplasma encephalomyelitis in a lamb associated with a spinal cord anomaly. Br Vet J 1975;131:380-384. PMID: 1148804. 74. Hay WH, Shell LG, Lindsay DS, Dubey JP. Diagnosis and treatment of Neospora caninum infection in a dog. J Am Vet Med Assoc 1990;197:87-89. PMID: 2370226. 75. Helmick B, Otter A, McGarry J, Buxton D. Serological investigation of aborted sheep and pigs for infection by Neospora caninum. Res Vet Sci 2002;73:187. PMID: 12204640. 76. Hernandez J, Risco C, Donovan A. Association between exposure to Neospora caninum and milk production in dairy cows. J Am Vet Med Assoc 2001;219:632-635. PMID: 11549092. 77. Hill DE, Liddell S, Jenkins MC, Dubey JP. Specific detection of Neospora caninum oocyst in fecal samples from experimentally-infected dogs using the polymerase chain reaction. J Parasitol 2001;87:395-398. PMID: 11318571. 78. Huong LTT, Ljungstrom BL, Uggla A, Bjorkman C. Prevalence of antibodies to Neospora caninum and Toxoplasma gondii in cattle and water buffaloes in southern Vietnam. Vet Parasitol 1998;75:53-57. PMID: 9566094. 79. Innes EA, Lunden A, Esteban I, et al. A previous infection with Toxoplasma gondii does not protect against a challenge with Neospora caninum in pregnant sheep. Parasite Immunol 2001a;23:121-132. PMID: 11240903. 80. Innes EA, Wright SE, Maley S, et al. Protection against vertical transmission in bovine neosporosis. Int J Parasitol 2001b;31:1523-1534. PMID: 11595240. 81. Innes EA, Andrianarivo AG, Björkman C, Williams DJL, Conrad PA. Immune responses to Neospora caninum and prospects for vaccination. Trends Parasitol 2002;18:497-504. PMID: 12473366. 82. Jakubek EB, Bröjer C, Regnersen C, Uggla A, Schares G, Bjorkman C. Seroprevalences of Toxoplasma gondii and Neospora caninum in Swedish red foxes ( Vulpes vulpes). Vet Parasitol 2001;102:167-172. PMID: 11705663. 83. Jenkins M, Baszler T, Bjorkman C, Schares G, Williams D. Diagnosis and seroepidemiology of Neospora caninum-associated bovine abortion. Int J Parasitol 2002;32:631-636. PMID: 11943234. 84. Jolley WR, McAllister MM, McGuire AM, Wills RA. Repetitive abortion in Neospora-infected ewes. Vet Parasitol 1999;82:251-257. PMID: 10348105. 85. Kim JH, Sohn HJ, Hwang WS, et al. In vitro isolation and characterization of bovine Neospora caninum in Korea. Vet Parasitol 2000;90:147-154. PMID: 10828521. 86. Klein BU, Muller E. Seroprävalenz von antikörpern gegen Neospora caninum bei Hunden mit und ohne klinischem Neosporoseverdacht in Deutschland. Der praktische Tierarzt 2001;82:437-440.
87. Kobayashi Y, Yamada M, Omata Y, et al. Naturally-occurring Neospora caninum infection in an adult sheep and her twin fetuses. J Parasitol 2001;87:434-436. PMID: 11318580. 88. Koyama T, Kobayashi Y, Omata Y, et al. Isolation of Neospora caninum from the brain of a pregnant sheep. J Parasitol 2001;87:1486-1488. PMID: 11780846. 89. Landmann JK, Jillella D, O'Donoghue PJ, McGowan MR. Confirmation of the prevention of vertical transmission of Neospora caninum in cattle by the use of embryo transfer. Aust Vet J 2002;80:502-503. PMID: 12224621. 90. Lindsay DS, Rippey NS, Powe TA, Sartin EA, Dubey JP, Blagburn BL. Abortions, fetal death, and stillbirths in pregnant pygmy goats inoculated with tachyzoites of Neospora caninum. Am J Vet Res 1995;56:1176-1180. PMID: 7486395. 91. Lindsay DS, Kelly EJ, McKown R, et al. Prevalence of Neospora caninum and Toxoplasma gondii antibodies in coyotes ( Canis latrans) and experimental infections of coyotes with Neospora caninum. J Parasitol 1996a;82:657-659. PMID: 8691381. 92. Lindsay DS, Steinberg H, Dubielzig RR, et al. Central nervous system neosporosis in a foal. J Vet Diagn Invest 1996b;8:507-510. PMID: 8953546. 93. Lindsay DS, Dubey JP, Duncan RB. Confirmation that the dog is a definitive host for Neospora caninum. Vet Parasitol 1999a;82:327-333. PMID: 10384909. 94. Lindsay DS, Upton SJ, Dubey JP. A structural study of the Neospora caninum oocyst. Int J Parasitol 1999b;29:1521-1523. PMID: 10608437. 95. Lindsay DS, Dubey JP. Canine neosporosis. J Vet Parasitol 2000;14:1-11.
96. Lindsay DS, Ritter DM, Brake D. Oocyst excretion in dogs fed mouse brains containing tissue cysts of a cloned line of Neospora caninum. J Parasitol 2001a;87:909-911. PMID: 11534658. 97. Lindsay DS, Spencer J, Rupprecht CE, Blagburn BL. Prevalence of agglutinating antibodies to Neospora caninum in raccoons, Procyon lotor. J Parasitol 2001b;87:1197-1198. PMID: 11695399. 98. Lindsay DS, Weston JL, Little SE. Prevalence of antibodies to Neospora caninum and Toxoplasma gondii in gray foxes ( Urocyon cinereoargenteus) from South Carolina. Vet Parasitol 2001c;97:159-164. PMID: 11358632. 99. Lindsay DS, Little SE, Davidson WR. Prevalence of antibodies to Neospora caninum in white-tailed deer, Odocoileus virginianus, from the Southeastern United States. J Parasitol 2002;88:415-417. PMID: 12054027. 100. Magnino S, Vigo PG, Fabbi M. Isolation of a bovine Neospora from a newborn calf in Italy. Vet Rec 1999;144:456. PMID: 10343382. 101. Marsh AE, Barr BC, Madigan J, Lakritz J, Nordhausen R, Conrad PA. Neosporosis as a cause of equine protozoal myeloencephalitis. J Am Vet Med Assoc 1996;209:1907-1913. PMID: 8944807. 102. Marsh AE, Barr BC, Packham AE, Conrad PA. Description of a new Neospora species (Protozoa: Apicomplexa: Sarcocystidae). J Parasitol 1998;84:983-991. PMID: 9794642. 103. Marsh AE, Johnson PJ, Ramos-Vara J, Johnson GC. Characterization of a Sarcocystis neurona isolate from a Missouri horse with equine protozoal myeloencephalitis. Vet Parasitol 2001;95:143-154. PMID: 11223195. 104. McAllister MM, McGuire AM, Jolley WR, Lindsay DS, Trees AJ, Stobart RH. Experimental neosporosis in pregnant ewes and their offspring. Vet Parasitol 1996;33:647-655. 105. McAllister MM, Dubey JP, Lindsay DS, Jolley WR, Wills RA, McGuire AM. Dogs are definitive hosts of Neospora caninum. Int J Parasitol 1998;28:1473-1478. PMID: 9770635. 106. McAllister MM, Bjorkman C, Anderson-Sprecher R, Rogers DG. Evidence of point-source exposure to Neospora caninum and protective immunity in a herd of beef cows. J Am Vet Med Assoc 2000;217:881-887. PMID: 10997162. 107. McDole MG, Gay JM. Seroprevalence of antibodies against Neospora caninum in diagnostic equine serum samples and their possible association with fetal loss. Vet Parasitol 2002;105:257-260. PMID: 11934465. 108. Mineo TWP, Silva DAO, Costa GHN, et al. Detection of IgG antibodies to Neospora caninum and Toxoplasma gondii in dogs examined in veterinary hospital from Brazil. Vet Parasitol 2001;98:239-245. PMID: 11423182. 109. Miller CMD, Quinn HE, Windsor PA, Ellis JT. Characterization of the first Australian isolate of Neospora caninum from cattle. Aust Vet J 2002;80:620-625. PMID: 12465814. 110. Moore DP, Campero CM, Odeon AC, et al. Seroepidemiology of beef and dairy herds and fetal study of Neospora caninum in Argentina. Vet Parasitol 2002;107:303-316. PMID: 12163242. 111. Morales E, Trigo FJ, Ibarra F, Puente E, Santacruz M. Neosporosis in Mexican dairy herds: Lesions and immunohistochemical detection of Neospora caninum in fetuses. J Comp Pathol 2001;125:58-63. PMID: 11437517. 112. Nam HW, Kang SW, Choi WY. Antibody reaction of human anti- Toxoplasma gondii positive and negative sera with Neospora caninum antigens. Korean J Parasitol 1998;36:269-275. PMID: 9868893. 113. O'Handley R, Liddell S, Parker C, Jenkins M, Dubey JP. Experimental infection of sheep with Neospora caninum oocysts. J Parasitol 2002;88:1120-1123. PMID: 12537104. 114. Ooi HK, Huang CC, Yang CH, Lee SH. Serological survey and first finding of Neospora caninum in Taiwan, and the detection of its antibodies in various body fluids of cattle. Vet Parasitol 2000;90:47-55. PMID: 10828511. 115. Ordeix L, Lloret A, Fondevila D, Dubey JP, Ferrer L, Fondati A. Cutaneous neosporosis during treatment of pemphigus foliaceus in a dog. J Am Anim Hosp Assoc 2002;38:415-419. PMID: 12220024. 116. Otter A, Wilson BW, Scholes SFE, Jeffrey M, Helmick B, Trees AJ. Results of a survey to determine whether Neospora is a significant cause of ovine abortion in England and Wales. Vet Rec 1997;140:175-177. PMID: 9055394. 117. Paré J, Fecteau G, Fortin M, Marsolais G. Seroepidemiologic study of Neospora caninum in dairy herds. J Am Vet Med Assoc 1998;213:1595-1598. PMID: 9838960. 118. Patitucci AN, Phil M, Pérez MJ, Rozas MA, Israel KF. Neosporosis canina: presencia de anticuerpos séricos en poblaciones caninas rurales y urbanas de Chile. Arch Med Vet 2001;33:227-232. 119. Perlé KMD, Del Piero F, Carr RF, Harris C, Stromberg PC. Cutaneous neosporosis in two adult dogs on chronic immunosuppressive therapy. J Vet Diagn Invest 2001;13:252-255. PMID: 11482605. 120. Peters M, Wagner F, Schares G. Canine neosporosis: clinical and pathological findings and first isolation of Neospora caninum in Germany. Parasitol Res 2000;86:1-7. PMID: 10669128. 121. Peters M, Lutkefels E, Heckeroth AR, Schares G. Immunohistochemical and ultrastructural evidence for Neospora caninum tissue cysts in skeletal muscles of naturally infected dogs and cattle. Int J Parasitol 2001a;31:1144-1148. PMID: 11429181. 122. Peters M, Wohlsein P, Knieriem A, Schares G. Neospora caninum infection associated with stillbirths in captive antelopes ( Tragelaphus imberbis). Vet Parasitol 2001b;97:153-157. PMID: 11358631. 123. Petersen E, Lebech M, Jensen L, et al. Neospora caninum infection and repeated abortions in humans. Emerging Infectious Diseases 1999;5:278-280. PMID: 10221883. 124. Pipano E, Shkap V, Fish L, Savitsky I, Perl S, Orgad U. Susceptibility of Psammomys obesus and Meriones tristrami to tachyzoites of Neospora caninum. J Parasitol 2002;88:314-319. PMID: 12054004. 125. Pitel PH, Pronost S, Romand S, Thulliez P, Fortier G, Ballet JJ. Prevalence of antibodies to Neospora caninum in horses in France. Equine Vet J 2001;33:205-207. PMID: 11266072. 126. Pronost S, Pitel PH, Romand S, Thulliez P, Collobert C, Fortier G. Neospora caninum: premiére mise en évidence in France sur un avorton équin, analyse et perspectives. Prat Vét Equine 1999;31:31-34.
127. Reichel MP, Thorton RN, Morgan PL, Mills RJM, Schares G. Neosporosis in a pup. N Z Vet J 1998;46:106-110. PMID: 16032029. 128. Sager H, Fischer I, Furrer K, et al. A Swiss case-control study to assess Neospora caninum-associated bovine abortions by PCR, histopathology and serology. Vet Parasitol 2001;102:1-15. PMID: 11705647. 129. Sawada M, Park CH, Kondo H, et al. Serological survey of antibody to Neospora caninum in Japanese dogs. J Vet Med Sci 1998;60:853-854. PMID: 9713815. 130. Sawada M, Kondo H, Tomioka Y, et al. Isolation of Neospora caninum from the brain of a naturally infected adult dairy cow. Vet Parasitol 2000;90:247-252. PMID: 10842005. 131. Schares G, Heydorn AO, Cuppers A, Conraths FJ, Mehlhorn H. Hammondia heydorni-like oocysts shed by a naturally infected dog and Neospora caninum NC-1 cannot be distinguished. Parasitol Res 2001;87:808-816. PMID: 11688886. 132. Simpson VR, Monies RJ, Riley P, Cromey DS. Foxes and neosporosis. Vet Rec 1997;141:503. PMID: 9402726. 133. Šlapeta JR, Modry D, Kyselova I, Horejs R, Lukes J, Koudela B. Dog shedding oocysts of Neospora caninum: PCR diagnosis and molecular phylogenic approach. Vet Parasitol 2002;109:157-167. PMID: 12423929. 134. Söndgen P, Peters M, Bärwald A, et al. Bovine neosporosis: immunoblot improves foetal serology. Vet Parasitol 2001;102:279-290. PMID: 11731071. 135. Stenlund S, Bjorkman C, Holmdahl OJM, Kindahl H, Uggla A. Characterisation of a Swedish bovine isolate of Neospora caninum. Parasitol Res 1997;83:214-219. PMID: 9089715. 136. Tarantino C, Rossi G, Kramer LH, Perrucci S, Cringoli G, Macchioni G. Leishmania infantum and Neospora caninum simultaneous skin infection in a young dog in Italy. Vet Parasitol 2001;102:77-83. PMID: 11705654. 137. Thurmond MC, Anderson ML, Blanchard PC. Secular and seasonal trends of Neospora abortion in California dairy cows. J Parasitol 1995;81:364-367. PMID: 7776121. 138. Thurmond MC, Hietala SK. Culling associated with Neospora caninum infection in dairy cows. Am J Vet Res 1996;57:1559-1562. PMID: 8915429. 139. Thurmond MC, Hietala SK. Effect of Neospora caninum infection on milk production in first-lactation dairy cows. J Am Vet Med Assoc 1997;210:672-674. PMID: 9054999. 140. Thurmond MC, Hietala SK. Neospora caninum infection and abortion in cattle. 1999, 4. pp 425-431.
141. Tranas J, Heinzen RA, Weiss LM, McAllister MM. Serological evidence of human infection with the protozoan Neospora caninum. Clin Diagn Lab Immunol 1999;6:765-767. PMID: 10473533. 142. Trees AJ, Williams DJL. Neosporosis in the United Kingdom. Int J Parasitol 2000;30:891-893.
143. Trees AJ, Williams DJL. Vaccination against bovine neosporosis - the challenge is the challenge. J Parasitol 2003;89(Supplement):in press.
144. Vardeleon D, Marsh AE, Thorne JG, Loch W, Young R, Johnson PJ. Prevalence of Neospora hughesi and Sarcocystis neurona antibodies in horses from various geographical locations. Vet Parasitol 2001;95:273-282. PMID: 11223207. 145. Walsh CP, Duncan RB, Zajac AM, Blagburn BL, Lindsay DS. Neospora hughesi: experimental infections in mice, gerbils, and dogs. Vet Parasitol 2000;92:119-128. PMID: 10946135. 146. Williams JH, Espie I, van Wilpe E, Matthee A. Neosporosis in a white rhinoceros (Ceratotherium simum) calf. Tydskr S Afr Vet Ver 2002;73:38-43.
147. Wolfe A, Hogan S, Maguire D, et al. Red foxes ( Vulpes vulpes) in Ireland as hosts for parasites of potential zoonotic and veterinary significance. Vet Rec 2001;149:759-763. PMID: 11808662. 148. Woods LW, Anderson ML, Swift PK, Sverlow KW. Systemic neosporosis in a California black-tailed deer ( Odocoileus hemionus columbianus). J Vet Diagn Invest 1994;6:508-510. PMID: 7858040. 149. Wouda W, Moen AR, Visser IJR, van Knapen F. Bovine fetal neosporosis: a comparison of epizootic and sporadic abortion cases and different age classes with regard to lesion severity and immunohistochemical identification of organisms in brain, heart, and liver. J Vet Diagn Invest 1997;9:180-185. PMID: 9211238. 150. Wouda W. Neospora abortion in cattle, aspects of diagnosis and epidemiology. 1998, the Netherlands. Univ. Utrecht. pp 1-176 PhD Thesis.
151. Wouda W, Bartels CJM, Moen AR. Characteristics of Neospora caninum-associated abortion storms in dairy herds in the Netherlands (1995-1997). Theriogenology 1999a;52:233-245. PMID: 10734391. 152. Wouda W, Dijkstra T, Kramer AMH, van Maanen C, Brinkhof JMA. Seroepidemiological evidence for a relationship between Neospora caninum infections in dogs and cattle. Int J Parasitol 1999b;29:1677-1682. PMID: 10608454. 153. Yamane I, Kokuho T, Shimura K, et al. In vitro isolation and characterisation of a bovine Neospora species in Japan. Res Vet Sci 1997;63:77-80. PMID: 9368961.
Fig. 1
Life cycle of Neospora caninum. 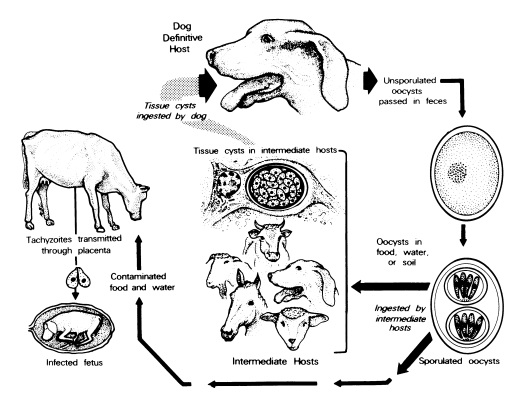
Fig. 2
Neospora caninum stages in dogs. Bar = 20 µm and applies to all figures. (A) Tachyzoites in an impression smear of lung. Giemsa stain. Note individual organisms (arrowheads) and those dividing into 2 (arrows). Compare size with red blood cells (rbc) and a macrophage (mo). (B) Tachyzoites in groups (arrows) and individuals (arrowheads) in sections of skin. Immunohistochemical stain with anti-N. caninum antibody. (C) Tissue cyst in section of brain. Note thick tissue cyst wall (arrow) enclosing bradyzoites (arrows). Toluidine blue stain. (D) Unsporulated oocyst with an individual sporont (arrow). Unstained. (E) Sporulated oocyst (arrow). Unstained. 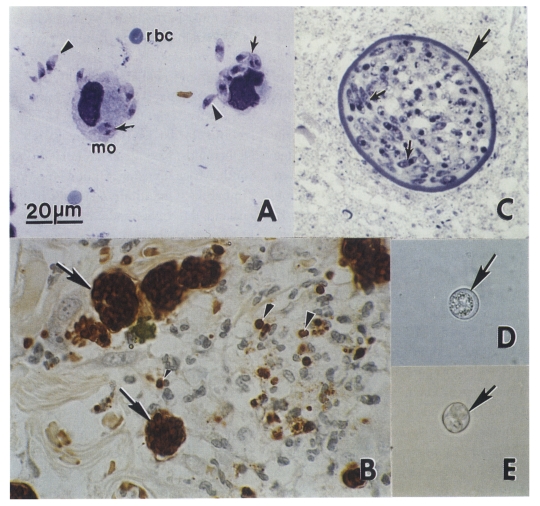
Table 1.
Diagnosis of N. caninum-associated abortion in cattle from selected studies
|
Country |
No. of fetuses examined |
Percent infected |
Reference |
|
Argentina |
240 |
12.1 |
Moore et al. (2002) |
|
Australia |
729 |
21.0 |
Boulton et al. (1995) |
|
Brazil |
46 |
39.1 |
Corbellini et al. (2002) |
|
Germany |
135 |
12.6 |
Sondgen et al. (2001) |
|
Mexico |
211 |
77 |
Morales et al. (2001) |
|
Switzerland |
242 |
21.0 |
Sager et al. (2001) |
|
The Netherlands |
2,053 |
17 |
Wouda et al. (1997) |
|
USA |
698 |
24.4 |
Anderson et al. (1991); Thurmond et al. (1995) |
|
USA |
266 |
42.5 |
Anderson et al. (1995) |
Table 2.
Neospora caninum isolates from cattle
|
Country |
Strain |
Source |
Reference |
|
Australia |
NC-Nowra |
7-day old calf |
Miller et al. (2002) |
|
Italy |
NC-PVI |
45-day old calf |
Magnino et al. (1999) |
|
Italy |
NC-PGI |
8-month old calf |
Fioretti et al. (2000) |
|
Japan |
JPA-1 |
Clinical calf |
Yamane et al. (1997) |
|
Japan |
BT-3 |
Adult cow |
Sawada et al. (2000) |
|
Korea |
KBA-1 |
1 day-old calf |
Kim et al. (2000) |
|
Korea |
KBA-2 |
Fetus |
Kim et al. (2000) |
|
Portugal |
NC-Porto1 |
Fetus |
Canada et al. (2002) |
|
Sweden |
NC-SweB1 |
Stillborn |
Stenlund et al. (1997) |
|
U.K |
NC-LivB1 |
Stillborn calf |
Davison et al. (1999) |
|
U.K |
NC-LivB2 |
Fetus |
Trees and Williams (2000) |
|
USA |
BPA-1 |
Fetus |
Conrad et al. (1993) |
|
USA |
BPA-2 |
Fetus |
Conrad et al. (1993) |
|
USA |
BPA-3 |
Clinical calf |
Barr et al. (1993) |
|
USA |
BPA-4 |
Clinical calf |
Barr et al. (1993) |
|
USA |
NC-Beef |
Clinical calf |
McAllister et al. (2000) |
|
USA |
NC-Illinois |
Clinical calf |
Gondim et al. (2002) |
Table 3.
Recognized isolates of Neospora caninum from dogs
|
Strain designation |
Country |
Reference |
|
NC-6- Argentinaa)
|
Argentina |
Basso et al. (2001a) |
|
NC-Bahiab)
|
Brazil |
Gondim et al. (2001) |
|
NC-GER1b)
|
Germany |
Peters et al. (2000) |
|
NC-livb)
|
UK |
Barber et al. (1995) |
|
NC-1b)
|
USA |
Dubey et al. (1988b) |
|
NC-2b)
|
USA |
Hay et al. (1990) |
|
NC-3b)
|
USA |
Cuddon et al. (1992) |
|
NC-4b)
|
USA |
Dubey et al. (1998a) |
|
NC-5b)
|
USA |
Dubey et al. (1998a) |
|
CN-1 |
USA |
Marsh et al. (1998) |
Table 4.
Seroprevalence of Neospora caninum antibodies in water buffaloes
|
Country |
No. of animals |
Type |
Testa)
|
Cut-off value |
% Positive |
Reference |
|
Brazil |
222 |
Dairy |
NAT |
1:40 |
53 |
Fujii et al. (2001) |
|
|
|
IFAT |
1:25 |
64 |
|
|
Egypt |
75 |
Meat |
NAT |
1:40 |
60 |
Dubey et al. (1998b) |
|
Italy |
1,377 |
Dairy |
IFAT |
1:200 |
34.6 |
Guarino et al. (2000) |
|
Vietnam |
200 |
Meat |
IFAT |
1:200 |
1.5 |
Huong et al. (1998) |
|
|
|
ELISA |
0.2 |
1.5 |
|
Table 5.
Seroprevalence of Neospora caninum antibodies in horses
|
Country |
No. of animals |
Test |
Cut-off value |
% Positive |
References |
|
Argentina |
76 |
NAT |
1:40 |
0 |
Dubey et al. (1999d) |
|
Brazil |
101 |
NAT |
1:40 |
0 |
Dubey et al. (1999c) |
|
France |
434 |
NAT |
1:40 |
23 |
Pitel et al. (2001) |
|
USA |
296 |
NAT |
1:40 |
21.3 |
Dubey et al. (1999b) |
|
536 |
IFAT |
1:50 |
11.5 |
Cheadle et al. (1999b) |
|
208 |
IFAT |
1:100 |
17 |
Vardeleon et al. (2001) |
|
300 |
IFAT |
1:50 |
10 |
McDole et al. (2002) |
|
South Korea |
191 |
IFAT |
1:100 |
2 |
Gupta et al. (2002) |
Table 6.
Seroprevalence of Neospora caninum antibodies in wildlife
|
Animal species |
Country |
No. examined |
Test |
% Positive |
Reference |
|
Red fox (Vulpes vulpes) |
Belgium |
123 |
IFAT |
78 |
Buxton et al. (1997a) |
|
Ireland |
70 |
IFAT |
1.4 |
Wolfe et al. (2001) |
|
Spain |
122 |
PCR |
10.7 |
Almeira et al. (2002) |
|
Sweden |
221 |
ELISA |
0 |
Jakubek et al. (2001) |
|
UK |
52 |
IFAT |
2 |
Barber et al. (1997) |
|
UK |
16 |
IFAT |
6 |
Simpson et al. (1997) |
|
Gray foxes (Urocyon cinereoargeneteus) |
USA |
26 |
NAT |
15.4 |
Lindsay et al. (2001c) |
|
Coyotes (Canis latrans) |
USA |
52 |
IFAT |
10 |
Lindsay et al. (1996a) |
|
White-tailed deer (Odocoileus virginianus) |
USA |
400 |
NAT |
40.5 |
Dubey et al. (1999a) |
|
|
305 |
NAT |
48 |
Lindsay et al. (2002) |
|
Australian dingos (Canis familiaris dingo) |
Australia |
169 |
IFAT |
15.9 |
Barber et al. (1997) |
|
Chiloe fox (Pseudolapex fulvipes) |
|
2 |
IFAT |
100 |
Patitucci et al. (2001) |
|
Lions (Panthera leo) |
S. Africa |
18 |
IFAT |
16.6 |
Cheadle et al. (1999a) |
|
Cheetah (Acinonyx jubatus) |
S. Africa |
16 |
IFAT |
6.3 |
Cheadle et al. (1999a) |
|
Raccoons (Procyon lotor) |
USA |
95 |
NAT |
10 |
Lindsay et al. (2001b) |
|
|





