INTRODUCTION
Strains and isolates of Toxoplasma gondii exhibit virulence differences (Suzuki et al., 1989; Sibley and Boothroyd, 1992). Virulent strains are those with a LD100 (100% lethal dose) of < 10 tachyzoites in outbred Swiss mice, whereas avirulent strains have a LD100 of > 103 tachyzoites (Sibley and Boothroyd, 1992). Therefore, one would expect genetic diversity among strains of T. gondii (Lehmann et al., 2000; Dubey et al., 2002). Recently, T. gondii strains were classified into three genotypes (I, II and III) by restriction fragment length polymorphism (RFLP) analysis (How and Sibley, 1995; Howe et al., 1997). All three genotypes can cause human infections; however, the genotype II predominates in human toxoplasmosis and in AIDS patients (Howe and Sibley, 1995; Howe et al., 1997; Grigg et al., 2001b). Moreover, molecular analyses of different geographical strains have led to hypothesize that the genus Toxoplasma contains more than one species, having distinct life cycles (Johnson, 1998). In this respect, isolation and characterization of as many geographical strains as possible are needed to properly evaluate this new phylogenic problem.
Tachyzoites of T. gondii were isolated from the blood of an ocular patient, and were successfully maintained in the laboratory for the first time in the Republic of Korea; this isolate was designated KI-1 (Chai et al., 2003). KI-1 was proved highly virulent when inoculated into BALB/c mice and cultured in sarcoma 180 cells (Chai et al., 2003). However, its genotype and other genetic characteristics were unknown. In the present study, PCR-RFLP patterns of dense granule antigen GRA6 and surface antigen SAG2 genes were used to determine the genotype of KI-1. In addition, the coding regions of the surface antigen gene SAG1, rhoptry antigen gene ROP1, and of granule antigen gene GRA8 were sequenced, and compared with those of RH strain.
MATERIALS AND METHODS
RNA extraction of T. gondii tachyzoites
Tachyzoites of T. gondii (KI-1 and RH) were collected from the peritoneal cavity of BALB/c mice and purified using 40% Percoll (Pharmacia Biotech, Uppsala, Sweden) in phosphate-buffered saline (PBS). Total RNA was extracted using a mono-phasic solution of phenol and guanidine isothiocyanate (Trizol® Reagent, Invitrogen, Carlsbad, CA, U.S.A.). Briefly, tachyzoites were homogenized in 1 ml of Trizol Reagent in a test tube on ice. Insoluble material was removed by centrifugation at 12,000 × g for 10 min at 4℃ and the supernatant was collected and incubated at 15-30℃ for 5 min. Chloroform (20% on total volume) was then added, the tube was shaken vigorously for 15 sec, incubated at 15-30℃ for 2-3 min and centrifuged at 12,000 × g for 15 min at 4℃. The upper layer was transferred to another tube, isoprophyl alcohol (33% on total volume) added and mixed and the tube incubated at room temperature for 10 min. The tube was then centrifuged at 12,000 × g for 10 min at 4℃, and the sediment re-suspended in 75% ethanol. The tube was vortexed and centrifuged at 7,500 × g for 5 min at 4℃. The pellet was air-dried, dissolved in an appropriate volume of 0.1% diethylpyrocarbonate solution and stored at -70℃.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
The cDNA of T. gondii RH and KI-1 was prepared by a first strand cDNA synthesis kit (AMV, Roche, Mannheim, Germany). Briefly, the reaction buffer containing 25 mM MgCl2, deoxynucleotide mix, RNase inhibitor, and reverse transcriptase were mixed in a sterile microfuge tube to produce a Master Mix. This sample was divided into 6 tubes; oligo-p(dT)15 primer was used in 3 tubes and random primer p(dN)6 in the remainder (one each for KI-1 and RH RNA, and a positive control). The tubes were then briefly vortexed, centrifuged, and incubated at 25℃ for 10 min and then at 42℃ for 60 min. AMV reverse transcriptase was then denatured by incubating the tube at 99℃ for 5 min and then cooling to 4℃ for 5 min. The products of the first strand cDNA were determined by agarose gel electrophoresis.
Regions encoding the GRA6 gene were amplified as previously described (Fazaeli et al., 2000a). The forward and reverse primers were 5'-GTAGCGTGCTTGTTGGCGAC-3' and 5'-TACAAGACATAGAGTGCCCC-3', respectively. PCR amplification was performed using the following conditions; 1 cycle of initial denaturation at 95℃ for 5 min followed by 30 amplification cycles (95℃ for 1 min, 60℃ for 1 min, and 72℃ for 1 min) and 1 cycle of final extension at 72℃ for 10 min. Amplification was carried out in a DNA thermal cycler (PE Applied Biosystems, Foster, California, U.S.A.).
PCR-based restriction fragment length polymorphism (PCR-RFLP) analysis
The PCR products were examined by 1.6% agarose gel electrophoresis and ethidium bromide staining. Amplified fragments were purified using a QIAGEN® II gel extraction kit (QIAGEN GmbH, Hilden, Germany) and digested with Mse I endonuclease. Restriction fragments were analyzed by agarose gel electrophoresis. SAG2 locus genotype was analyzed using a PCR approach that separately amplified the 5' and 3' ends of the locus, as described previously (Howe et al., 1997). Briefly, the 5' end of the locus was amplified by 40 standard PCR cycles using primers F1 (5'-GAAATGTTTCAGGTTGCTGC-3') and R1 (5'-GCAAGAGCGAACTTGAACAC-3'). The 3' end of the locus was similarly amplified using primers F2 (5'-ATTCTCATGCCTCCGCTTC-3') and R2 (5'-AACGTTTCACGAAGGCACAC-3'). The amplification products were purified using a gel extraction kit (QIAGEN) and digested with Sau3A I and Hha I, respectively. Restriction fragments were then analyzed by agarose gel electrophoresis.
Sequencing of the SAG1, ROP1, and GRA8 genes
The SAG1, ROP1, and GRA8 genes of RH and KI-1 were amplified, using the primers made in this study; F (ACGAATTCGACGAGTATGTTT) and R (ACGAATTCAACGGTGATC) for SAG1, F (ATGGAGCAAAGGCTG CCAAT) and R (TTATTGCGATCCATCATCCTG) for ROP1, and F (ATGGCTTTACCATTGCGTG) and R (TTAATTCTGCGTCGTTA CGG) for GRA8. The PCR products were purified using a QIAGEN® II gel extraction kit (QIAGEN), and sequenced on an automated sequencer (Models 373A and 377, PE Applied Biosystems) by the dideoxy-mediated chain termination method. The primers for PCR amplification were also used for the sequencing reactions. Alignments and comparisons of nucleotide and amino acid sequences were performed using Lagergene biocomputing (DNASTAR Inc., version 2.1, Madison, Wisconsin, U.S.A.) and Sequence Navigator (PE Applied Biosystems) software. Nucleotide sequence data obtained in this study have been deposited in the GenBank (Bethesda, Maryland, U.S.A.), under the accession numbers AY661790 (ROP1) and AY661791 (SAG1).
RESULTS
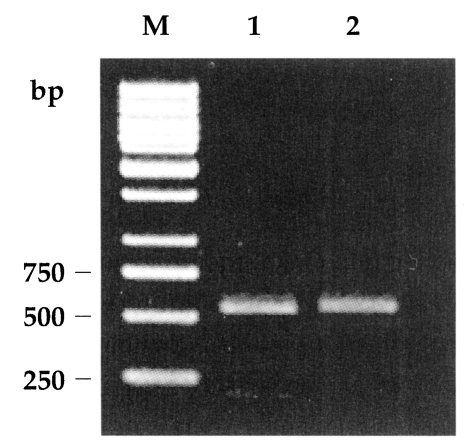
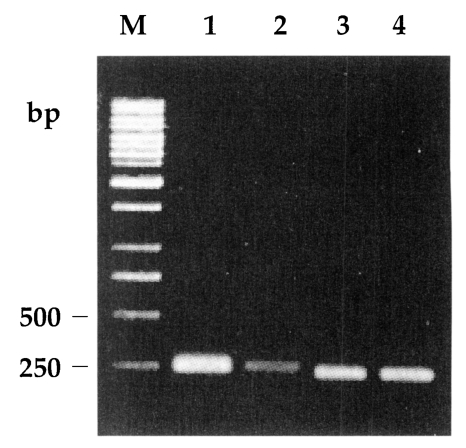
Amplification of the GRA6 gene yielded a 800-bp product for both RH and KI-1. Digestion of the amplification products of RH and KI-1 with Mse I endonuclease resulted in two bands at 195 and 544 bp (Fig. 1), which was consistent with the genotype I of T. gondii (Fazaeli et al., 2000a). The 5' and 3' ends of the SAG2 locus were successfully amplified and resulted in 241-bp and 221-bp products, respectively (Fig. 2). Meanwhile, no Sau3A I or Hha I endonuclease sites existed in the 5' or 3' ends of SAG2 of both RH and KI-1 (Fig. 2), not different from the genotype I (Howe et al., 1997).
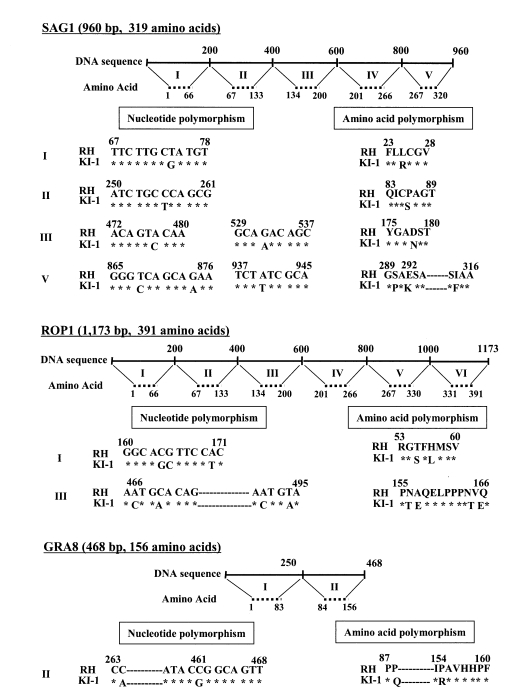
The alignment of the region encoding the SAG1 gene in KI-1 showed nucleotide substitutions at 7 positions (about 1% of all positions), versus SAG1 in RH (Fig. 3). This resulted in amino acid substitutions at 6 positions. Seven nucleotide substitutions were also observed in the ROP1 gene coding region of KI-1 (Fig. 3), versus RH, and 6 of the 7 substitutions resulted in amino acid sequence changes. The nucleotide sequence alignment of KI-1 and RH also showed nucleotide polymorphisms in GRA8 gene at 2 positions (about 1% of all positions), respectively (Fig. 3), which all resulted in amino acid substitutions. However, no deletions or insertions were found in the SAG2, ROP1, or GRA8 genes of KI-1, compared with those of RH.
DISCUSSION
By PCR-RFLP analyses (Howe and Sibley, 1995; Howe et al., 1997), 3 clonal lineages were identified among T. gondii strains and isolates. The genotype I is virulent, whereas the genotype II is relatively avirulent. On the other hand, the genotype III consists of avirulent and less virulent isolates (Howe and Sibley, 1995), and has been shown to possess a combination of virulent and avirulent alleles (Grigg et al., 2001a). However, not all virulent strains produce the same RFLP patterns. For example, MAS strain, which is highly pathogenic to mice, was typed as the genotype I using 6 different genetic loci. However, different patterns were observed in 2 of the 6 loci studied (Howe and Sibley, 1995). We previously reported that KI-1 is virulent (Chai et al., 2003), and expected to be of the genotype I clonal lineage. In the present study, the genotype of KI-1 was determined by analyzing the 5' and 3' ends of the SAG2 locus and GRA6 locus because these genes contain multiple lineage-specific polymorphisms and are ideally suited for rapid genotyping using PCR-RFLP (Howe et al., 1997; Fazaeli et al., 2000a). The results revealed that KI-1 is a member of the genotype I (How et al., 1997; Fazaeli et al., 2000a).
Gene sequence analysis has been performed in a number of DNA loci of different T. gondii strains, including the heat-shock protein 70 gene (Lyons and Johnson, 1998) and the intergenic spacer gene (Fazaeli et al., 2000b). Sequence variants of these genes have been used as markers to distinguish T. gondii isolates from different hosts and geographical localities (Høgdall et al., 2000). In the present study, sequencing was performed on the encoding regions of SAG1, ROP1, and GRA8 genes for RH and KI-1; the results showed nucleotide and amino acid polymorphisms of these genes at about 1% and 2% of the overall sequences, respectively. Although further studies are required to clarify the significance of these differences, at present we regard them as minor variations that can occur between geographical strains of the same genotype groups.
It is interesting to note that most human toxoplasmosis is caused by the genotype II, and only a small proportion by genotypes I and III (Howe and Sibley, 1995; Howe et al., 1997). Genotypes I and II occur more frequently in AIDS and congenitally infected patients than in animals, whereas the genotype III predominates in animals (Howe and Sibley, 1995). However, in the case of severe ocular diseases in man, strains of the genotype I and recombinants of genotypes I and III are more common than the genotype II (Grigg et al., 2001b). KI-1 originated from a patient with severe ocular disease (Chai et al., 2003); therefore, the results of the present study agree with those of a previous report (Grigg et al., 2001b).
Conclusively, based on genotypic properties, KI-1 is included in the clonal lineage of the genotype I, which also includes RH. KI-1 revealed minor nucleotide and amino acid substitutions, and could be considered a new geographical strain.