INTRODUCTION
Acanthamoeba spp. are ubiquitous free-living amoebae which function as predators that control microbial communities; these species have also been recognized as human pathogens that cause keratitis or granulomatous amebic encephalitis [1]. In addition, a variety of other clinically-related bacteria, including Mycobacterium avium, Vibrio cholerae, Listeria monocytogenes, Legionella pneumophila, and Burkholderia cepacia, are capable of surviving and multiplying for a limited time within acanthamoebae [2-6]. It has been suggested that growth in an amebic intracellular environment might help these bacteria to adapt and survive within mammalian phagocytic cells [7-9]. Moreover, the endosymbiosis of bacteria in amoebic cysts has also been demonstrated to be relatively resistant to harmful environmental conditions, including exposure to antibiotic agents [10,11].
Several studies concerning the long-term symbiotic relationship between Acanthamoeba and bacteria have been conducted since the first such study in the 1970's. These obligate bacterial endosymbionts have been reported in approximately 24% of all Acanthamoeba isolates [12]. However, the majority of obligate endosymbiotic bacteria could not be cultured via standard microbiological techniques. Recently, many researchers have attempted to characterize the diversity and phylogeny of these obligate intracellular microorganisms via molecular identification techniques. In some studies, comparative analyses of 16S rRNA sequences have successfully demonstrated the presence of several endosymbionts in Acanthamoeba, including Legionella [13], Chlamydiales [14], Rickettsiales, or Caedibacter-related bacteria [15,16].
In a previous study, we detected Acanthamoeba contamination in domestic tap water, and evaluated molecular characteristics of these specimens using 18S rDNA sequencing and mitochondrial (Mt) DNA RFLP techniques [17]. We determined that some Acanthamoeba strains harbored endosymbiotic bacteria, via characterization by Mt DNA RFLP. In this study, we have confirmed the presence of endosymbiotic bacteria, using transmission electron microscopy. In order to estimate their pathogenicity, we conducted a genetic characterization of the endosymbionts in Acanthamoeba isolates from domestic tap water using 16S rDNA sequencing.
MATERIALS AND METHODS
Orcein staining
Orcein staining was utilized to visualize the presence of endosymbionts within the Acanthamoeba isolates, as described by Beier et al. [15]. Thin films of Acanthamoeba were prepared on slides and allowed to dry in air. The slides were placed in 1% KMnO4 for 5 min, washed with water, then rinsed for 30 sec in 3% oxalic acid and finally, washed off in distilled water. The films were covered with a few drops of the orcein-carmine stain solution. The slides were subsequently examined 1 hr after staining, under a light microscope (Olympus IX70, Tokyo, Japan) using oil immersion objectives.
Transmission electron microscopy
Trophozoites were washed 3 times with cold PBS and then pre-fixed for 2 hr with 4% glutaraldehyde in 0.1 M cacodylate buffer. After rinsing with 0.1 M cacodylate buffer, the sediment was post-fixed for 3 hr with 1% osmium tetroxide, rinsed twice with 0.1 M maleate buffer at pH 5.2, dehydrated with ethyl alcohol, and treated for 30 min with propylene oxide. The pellet was immersed overnight in propylene oxide-resin (1 : 1) with continuous shaking, and then embedded in resin and incubated overnight at 60℃. Ultrathin sections cut on a Reichert-Jung ultramicrotome were stained with uranyl acetate and lead citrate. The sections were observed under a transmission electron microscope (Hitachi H-7000, Tokyo, Japan).
Sequence analysis of 16S rDNA of endosymbionts and 18S rDNA of Acanthamoeba
In order to amplify the 16S rDNA of bacterial endosymbionts, the Mt DNA of each Acanthamoeba was utilized as a template. The nucleotide sequences of the primers were as follows: forward 5'-TCGACAACAGAGTTTGATCCTGGCTCAG-3', reverse 5'-ATCCAAGCTTAAGGAGGTGATCCAGCC-3'. The 16S rDNA amplification reactions were conducted separately in a thermal capillary cycler using premix of 50 µl scale (Bioneer, Seoul, Korea), including 1.25 U Taq polymerase (Bioneer). The PCR reactions were conducted for an initial 3 min at 94℃ followed by 30 cycles of 94℃ for 1.5 min, 50℃ for 1 min, 72℃ for 4 min, and a final elongation step of 7 min at 72℃. Full-length amoeba 18S rDNA genes were amplified and sequenced as previously reported [20]. In brief, the PCR reactions were conducted for an initial 3 min at 94℃ followed by 30 cycles of 94℃ for 1 min, 58℃ for 1 min, 72℃ for 2.5 min, and a final 10 min elongation step at 72℃. The amplified PCR products were ligated into the T/A cloning vector pGEM-T easy System I (Promega, Madison, Wisconsin, USA), with subsequent transformation in E. coli by each vector. DNA sequencing was performed by an outside company (MACROGEN, Seoul, Korea).
Phylogenetic analyses
Nucleotide sequences were compared to those of published strains via BLAST searches [21]. Clustal X [22] was used to conduct pairwise alignment and to calculate the percentage of sequence dissimilarity. The percent similarity corresponds to the number of identical sites between the 2 isolates, aligned to each other as in the master alignment. Phylogeny was determined using the ClustalW2 program (http://www.ebi.ac.uk/Tools/clustalw2/index.html) with a low gap penalty, and phylogenetic trees were constructed with TreeView computer software.
RESULTS
Endosymbionts in Acanthamoeba detected by orcein staining
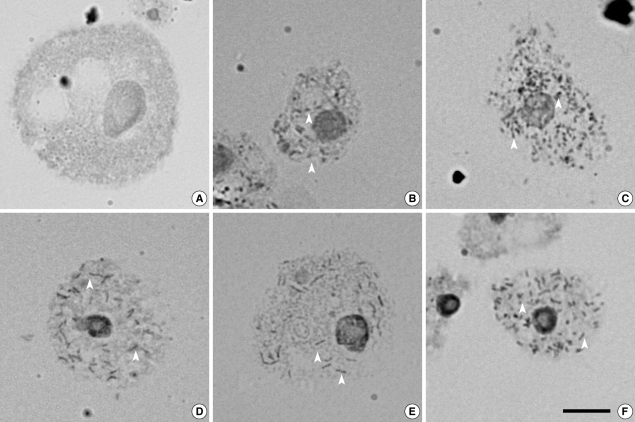
Five isolates (29.4%) of Acanthamoeba were found to harbor endosymbionts in their cytoplasm. The bacterial endosymbionts were readily visualized within Acanthamoeba stained with orcein under phase-contrast light microscopy (Fig. 1). Darkly stained rod-shaped bacterial endosymbionts were distributed randomly in the cytoplasm of trophozoites of Acanthamoeba sp. KA/WP3, KA/WP4, KA/WP8, KA/WP9, and KA/WP12. In comparison, no bacteria were detected in A. castellanii Castellani, which was employed as a reference strain.
Transmission electron microscopy
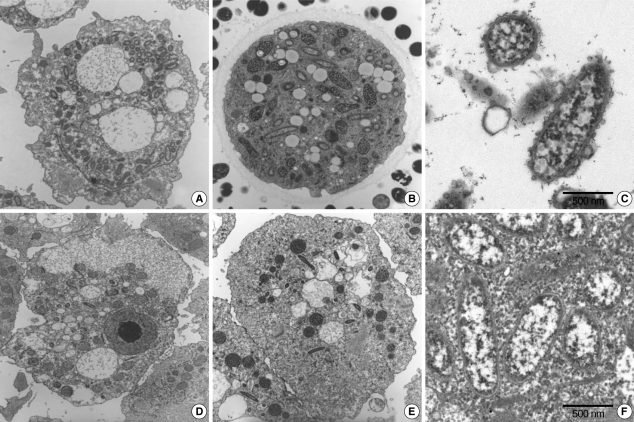
The morphological and ultrastructural characters of the endosymbionts were elucidated via transmission electron microscopy. We found that it was substantially easier to detect endosymbionts in the ameba than in A. castellanii Castellani. The rodshaped endosymbionts detected in KA/WP3 and KA/WP4 were estimated to be approximately 1.72 (1.70-1.75) × 0.27 (0.20-0.30) µm in size (Fig. 2A-C), and the endosymbionts (1.55 × 0.25 µm) detected in KA/WP8 and KA/WP12 were smaller than those observed in KA/WP3 and KA/WP4 (Fig. 2D-F). The endosymbionts of KA/WP3 and KA/WP4 were quite similar to each other in terms of morphological characteristics, and they could be readily detected in the cytoplasm of Acanthamoeba, because they existed at numbers in excess of 500 per Acanthamoeba. Additionally, we noted that these endosymbionts were bound by double membranes and their cell surfaces were studded with numerous host cell ribosomes (Fig. 2C).
We also noted some different morphological characteristics in the endosymbionts found in KA/WP8 and KA/WP12; the endosymbionts in these strains were bound by double membranes, but also exhibited apparent vesicle-like structures on the bacterial outer membrane; a translucent zone surrounding the endosymbiont was also observed (Fig. 2F). Additionally, the endosymbionts of KA/WP3, KA/WP4, KA/WP8, and KA/WP12 were found to be distributed randomly throughout the cytoplasm of the trophozoites and cyst forms, but the endosymbionts of KA/WP9 were distributed with group (2-16 endosymbionts per group) in the cytoplasm (Fig. 3). However, these endosymbionts are not surrounded by any membranes or vesicles. The endosymbionts of KA/WP9 also evidenced a very different shape than the other endosymbionts in this study; their bodies were curved rod-shaped and quite irregular (asymmetry rod shape).
16S rRNA sequences of endosymbionts
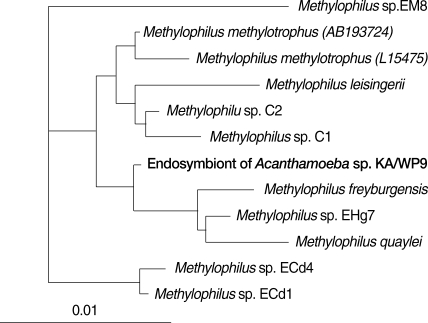
Comparative analysis with 16S rRNA gene sequences demonstrated that the endosymbionts of KA/WP3 and KA/WP4 evidenced 99% sequence similarity with each other. The endosymbionts of KA/WP3 and KA/WP4 showed the highest similarity, at 97% of the recently published 16S rDNA sequence of the bacteria, Candidatus Amoebophilus asiaticus [23]. Additionally, these symbionts evidenced a high degree of similarity with the endosymbionts of the tick, Ixodes scapularis (89% and 90%, respectively). The phylogenetic analysis consistently demonstrated that the endosymbiont of KA/WP3 and KA/WP4, Candidatus Amoebophilus asiaticus, and the endosymbiont of I. scapularis form a monophyletic lineage within the CFB phylum (Fig. 4). However, the endosymbiont of KA/WP3 and KA/ WP4 demonstrated only moderate similarity (less than 80%) with the other endosymbionts detected in KA/WP8, KAWP9, and KAWP12. Comparative analysis with 16S rRNA gene sequences showed that the endosymbionts of KA/WP8 and KA/WP12 shared 97% sequence similarity with each other. These endosymbionts evidenced a high degree of similarity to Candidatus Odyssella thessalonicensis, a member of the α-proteobacteria. They share an evolutionary line of descent with Caedibacter caryophilus. The endosymbiont of KA/WP9 is highly similar (85-95%) to genus Methylophilus, which has not previously been determined to harbor any endosymbionts (Fig. 5). This is the first report to show that Methylophilus spp. can live in the cytoplasm of Acanthamoeba.
18S rRNA sequences of 3 Acanthamoeba strains
The dendrogram constructed on the basis of the 18S rDNA sequence analyses is shown in Fig. 6. Comparative sequence analysis verified the morphology-based identification of this isolate as Acanthamoeba sp. KA/WP3 evidenced the highest level of sequence similarity (98%) with A. castellanii Castellani. In addition, Acanthamoeba KA/WP3 evidenced a high 18S rDNA sequence similarity (99%) with the clinical isolates of KA/E1 (isolated from Korean patients with keratitis). Acanthamoeba KA/WP4, KA/WP8, and KA/WP12 exhibited the highest level of sequence similarity (98%) with A. lugdunensis L3a. These amebas also exhibited very high 18S rDNA sequence similarity (97%) with the clinical isolates KA/E2, KA/E12, KA/E15. Acanthamoeba KA/WP9 is closely related to A. rhysodes Singh, which was isolated from soil. This amoeba also has not previously been recognized as a host for any endosymbionts. Using the 95% similarity value for the definition of the Acanthameoba 18S rDNA sequence types [24], all of the host Acanthamoeba in this study could be assigned to genotype T4, which comprises the majority of clinical and environmental Acanthamoeba isolates.
DISCUSSION
Bacterial endosymbionts are commonly observed in protozoans, particularly for Acanthamoeba; approximately 24% of Acanthamoeba isolates from both environmental and medical samples were determined to harbor endosymbionts [12,25]. In our study, 29.4% of Acanthamoeba isolated from domestic tap water harbored endosymbiont bacteria in their cytoplasm.
A number of endosymbionts of both KA/WP3 and KA/WP4 were detected in the cytoplasm of Acanthamoeba, and they were identified in this study as Candidatus Amoebophilus asiaticus, via 16S rDNA sequencing analysis. This bacteria was initially described by Horn et al. [23], who explained that "Amoebophilus" taxonomically designates the obligate intracellular lifestyle of the bacteria in free-living amoebae, and the term "asiaticus" refers to continental Asia, where the host was originally isolate. Two independent laboratories previously attempted to culture the bacteria in a number of cell-free media, but they failed [23]. These findings suggested that eukaryotic host cells are essential for the growth of these bacteria, similarly to mitochondria. We also speculate that the endosymbiont did not affect, or only minimally affected the biological functions of the ameba, because a number of bacteria (especially endosymbionts of KA/WP3) occupied the ameba cytoplasm although there were no pathological findings (Fig. 2A). These endosymbionts were surrounded by a double membrane decorated with numerous ribosomes of the host Acanthamoeba. These morphological characteristics are consistent with those reported for the endosymbionts of Acanthamoeba isolated previously from contact lens storage cases in Korea and other countries [26-28].
We also identified other obligate intracellular bacteria detected in KA/WP8 and KA/WP12 as Candidatus Odyssella thessalonicensis. This species name was suggested by Birtle et al. [28], and referenced Odysseus and Thessalonika (the Greek city). These bacteria cannot be cultivated on cell-free medium, and can only be co-cultivated with a number of Acanthamoeba species at temperatures at or below 37℃. At temperatures above 30℃, the organism multiplies and rapidly destroys its host [28]. We noted an electron-translucent area surrounding the bacteria, which was attributed to the presence of bacterial capsular material. Similar endosymbionts were also reported [28,29].
In this study, we identified an endosymbiont from KA/WP9, the morphological and molecular characteristics of which were quite similar to those of Methylophilus spp. Methylophilus was suggested as a new genus by Jankins et al. in 1987; they described the characterization of the bacteria as follows [30]. When grown on methanol-mineral salts agar or in methanol mineral salts liquid medium, straight, or slightly curved rods usually 0.3 to 0.6 pm in diameter and 0.8 to 1.5 pm in length occur either singly or in pairs; they are gram-negative, but the stain is frequently not taken up well. They are motile (by polar flagella) or non-motile. Endospores are absent; no cellular inclusions; no sheath or prosthecae detected; no capsules formed, but slime may be produced by some strains [30]. The bacteria occur singly or in pairs and can be isolated from sludge, mud, and river and pond water [31]. Because their niche is shared with Acanthamoeba, they can be fed by Acanthamoeba. However, until now, Methylophilus has not previously been recognized as an endosymbiont occupying Acanthamoeba.
We identified endosymbionts (Methylophilus spp.) from KA/WP9 using electron microscopy after 6 months from axenic culture. We could check for endosymbionts from KA/WP9 until 1 year after identification. However, the KA/WP9 culture medium became contaminated by unknown bacteria, and thus we replaced the culture medium with one containing antibiotics. Afterward, we were unable to detect any endosymbionts in KA/WP9. We are still uncertain as to whether the contaminating bacteria were endosymbionts or not. The endosymbionts of KA/WP9 was not surrounded by host phagocytic membranes (Fig. 3F), and only a few bacteria were detected in the cytoplasm of KA/WP9 (Fig. 3E). We believe that this stage may be an initial step in the symbiosis between the amoeba and the bacteria.
Many medically important bacteria are capable of utilizing Acanthamoeba as shelter; in particular, many studies have focused on Legionella and Mycobacterium in this regard [32,33]. We don't know whether or not the endosymbiont of KA/WP9, Methylophilus, is virulent, because we failed in our attempts to cultivate these bacteria with Methylophilus-specific medium. With the exception of the presence of endosymbionts, Acanthamoeba sp. KA/WP3 and KA/WP4 with Acanthamoeba sp. KA/E1 (keratitis isolates), as well as KA/WP8 and KA/WP12 with A. lugdunensis L3a and Acanthamoeba sp. KA/E2 were phylogenetically closely related, according to the results of Mt DNA RFLP and 18S rDNA sequence analysis [17]. They are most frequently isolated from contact lens storage cases and keratitis patients in Korea. This isogenetic pair can be utilized to evaluate the relationship of the bacteria with their host, Acanthamoeba. Further studies should focus on the functions of these endosymbionts, which enhance amoebic cytopathogenicity.