INTRODUCTION
Hepatic fibrosis occurs in the setting of chronic liver injury of different etiologies, including schistosomiasis. Schistosomiasis is an important cause of hepatic fibrosis. Chronic infection by Schistosoma mansoni is one of the experimental models of hepatic fibrosis used to elucidate the mechanism involved in the fibrogenic process [1]. Thus, it is of great interest to explore new therapeutic strategies to ameliorate the damages caused by fibrogenesis in chronic liver diseases [2]. Several studies reported that hepatic fibrosis is a reversible disease; therefore, an effective treatment would probably prevent or reverse the fibrotic process in the liver [3,4].
The key event in S. mansoni infection is the formation of granulomas around eggs trapped in the portal venules of the liver tissue [5], and the eggs release a variety of substances, leading to antigen-specific humoral and cell-mediated immune responses [6] which is a complex pathophysiological cascade event which terminates in fibrosis and portal hypertension [7,8].
The granulomatous reaction results in a lesion 20 times larger than the egg itself. This reaction started with accumulation of T-cells which attract eosinophils and neutrophils around the living eggs to destroy the egg shell. Consequently, following initiation of the inflammation, macrophages secrete proinflammatory molecules such as IL-1, IL-6, and TNF-α [9]. Also, collagen is a group of naturally occurring proteins, and it is the main protein of connective tissues making about 25-35% of the whole body protein content [10]. Excessive production of collagen has been documented in proliferative disorders such as liver cirrhosis. In other clinical situations such as tissue repair and wound healing, the overproduction and deposition of collagen are required to heal the damaged tissues. In order to understand the critical role of collagen in various pathophysiological conditions, it is necessary to determine its content in liver tissues [11].
Stem cells were reported to have a potential role to support tissue regeneration, requiring minimally invasive procedures with few complications. They have recently shown promise in cell therapy because they have the capacity for self-renewal, multilineage differentiation, and are applicable to human diseases [12]. Transplanted BMSCs were reported to give rise to hepatocytes and exert significant influence on hepatic architecture according to Abdel Aziz et al. [13]. Also, Pai et al. [14] stated that transplantation of BMSCs can restore liver mass and function, alleviate fibrosis, and correct inherited liver diseases. Therefore, it can significantly improve the liver functions (albumin, ALT, AST, and ALP) of patients with terminal liver diseases with good safety and effectiveness. So, regenerative medicine has emerged as an alternative therapy to improve the damaged liver [15,16,17].
MATERIALS AND METHODS
Experimental animals
BALB/c mice were used for this experiment and bred at Theodor Bilharz Research Institute (TBRI), Egypt. Female mice were used as recipients while male mice were used as donors of BMSCs. All animal protocols were conducted in accordance with the valid international guidelines for animal experimentation and were approved by the TBRI's animal research committee.
Experimental design
A total of 96 female BALB/c mice were divided into 4 main groups (24 mice/group). Group A contained normal uninfected negative control mice which had injected intravenously with saline; the animals of group B were infected subcutaneously with S. mansoni cercariae (obtained from infected Biomphalaria alexandrina snails which were bred and maintained at the SBSP/TBRI) and not subjected to bone marrow stem cells inoculation. The animals of groups C and D were infected with S. mansoni cercariae and subjected to treatment with BMSCs (1 million cells/mouse) alone and BMSCs in combination with HGF, respectively on the 8th week post-infection (PI). BMSCs derived from male BALB/c mice were infused by peripheral injection route.
Eight animals from each group were sacrificed after the first, second, and third month post-treatment (PT) and their livers were subsequently processed for histological and immunohistological examinations. The liver samples were fixed in 10% buffered formalin for histopathological study. Blood samples were taken individually for sera preparation, and kept at -20℃ until used for examinations.
Isolation, culture, purification, and inoculation of BMSCs
Serum-free media were prepared using RPMI-1640, supplemented with L-glutamine, hepes buffer, sodium bicarbonate, and penicillin/streptomycin with pH 7.2-7.4. Then, complete media consisted of serum-free media supplemented with 20% fetal calf serum (FCS). BMSCs were harvested from the bone marrow of the femurs and tibiae of donor male mice according to the described method [18,19], with some modifications. Finally, the isolated cells were incubated for 48 hr at 37℃ under 5% CO2 for pre-stimulation of BMSCs. Cells viability was measured after 48 hr incubation and counted using a hemocytometer under graduated microsope. Then, a single intravenous inoculation of 1 million male unfractionated BMSCs was applied per mouse to the non-ablated female S. mansoni-infected mice of group C at week 8 PI. Also, another single inoculation with the same number of unfractionated BMSCs supplemented with HGF was injected intravenously into the non-ablated female S. mansoni-infected mice of group D at week 8 PI.
PCR detection of male derived BMSCs
The presence of donor-derived DNA in the liver tissue was analyzed after BMSCs transplantation in mice of all studied groups. The presence or absence of the sex determination region on the Y chromosome male (sry) gene in recipient female mice was assessed by PCR [18].
Hepatic histopathology and immunohistochemistry parameters
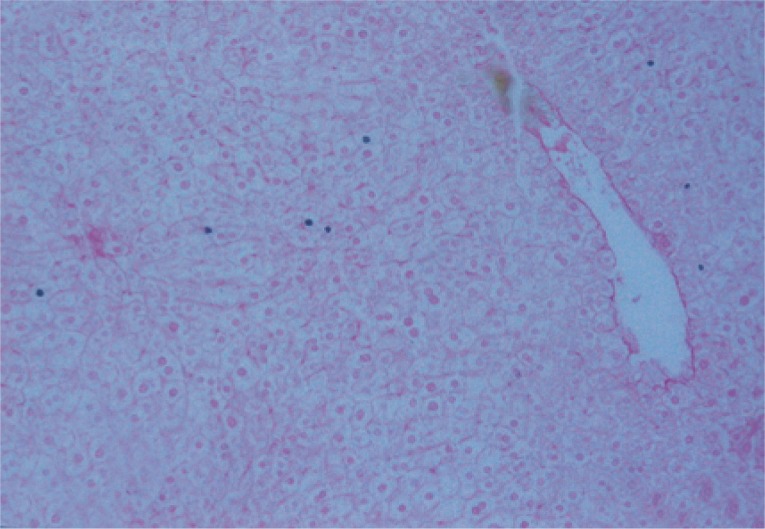
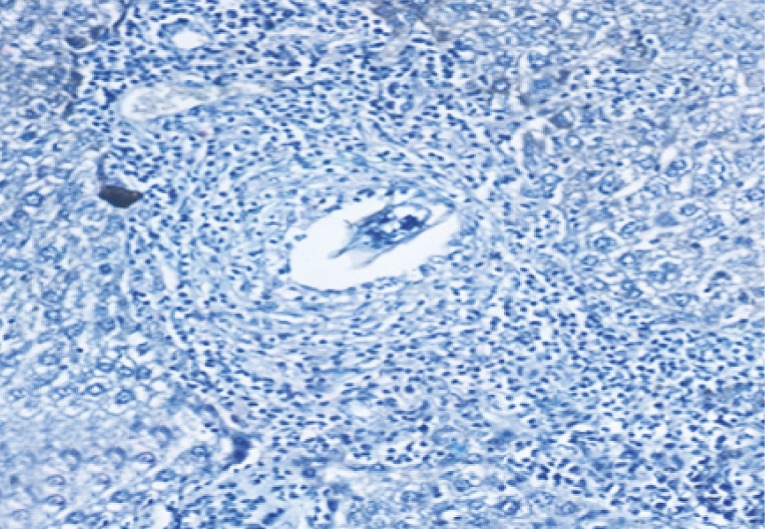
Liver specimens were fixed in 10% formalin, processed to paraffin blocks, and sectioned at 4 µm thickness. Sections were stained with hematoxylin and eosin (H&E) to study the hepatic morphological changes and hepatic schistosomal fibrosis [19].
Liver sections were examined by a light microscope regarding the exerted hepatic schistosomal infection and portal fibrosis through the periods of the experiment [20]. This study focused on tracing the homing of the transplanted BMSCs and their differentiation into newly formed hepatocytes [19], applying immunohistochemistry techniques using monoclonal anti-mouse anti-OV-6 antibodies for detection of newly formed hepatocytes.
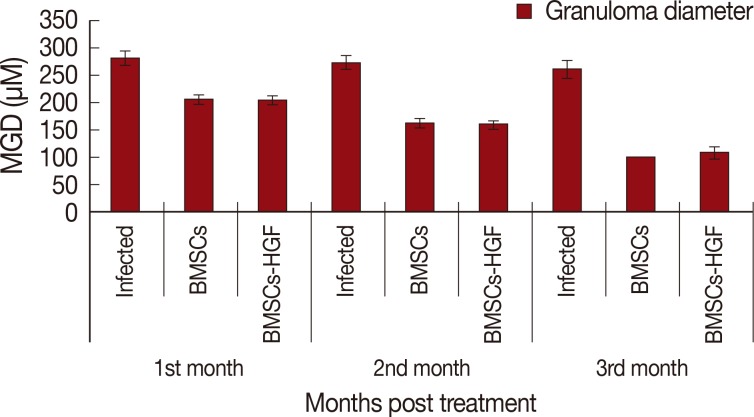
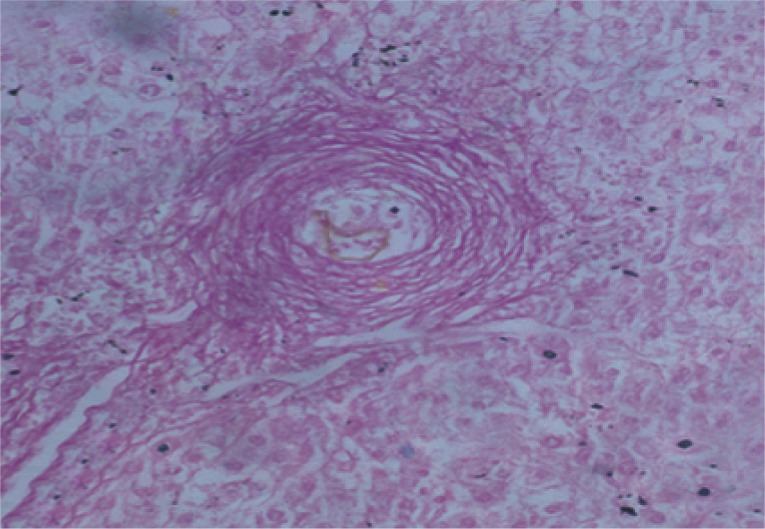
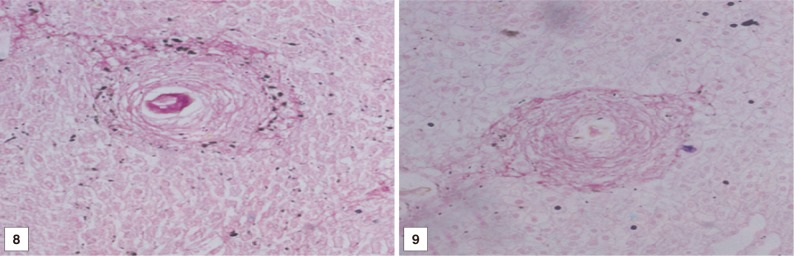
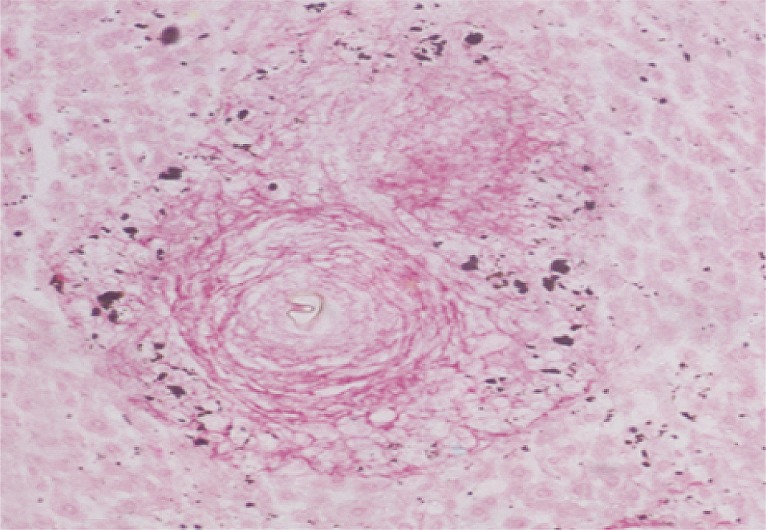
Measurement of granuloma diameter
Liver tissue lesions from 5, 6 animals were measured for each group. Measurements were done only for granulomas containing a single egg in their center. The mean granuloma diameter (MGD) was obtained in microns by measuring 2 diameters of the lesion at right angles to each other with the help of an ocular micrometer. The count was done under a light microscope. The MGD of each liver granuloma was obtained in microns by dividing the sum of the measured vertical and transverse diameters of the lesions by 2. The mean diameter of all lesions measured was then calculated for each group [21]. According to the method [22], lesion counts from 50 to 100 were taken into considerations. Then, the percent suppression of MGD/treated group was calculated.
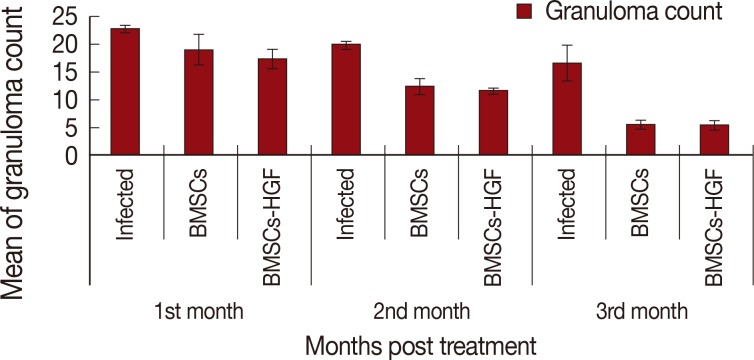
Measurement of granuloma number
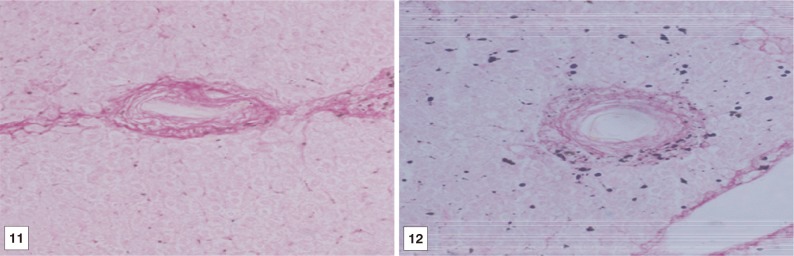
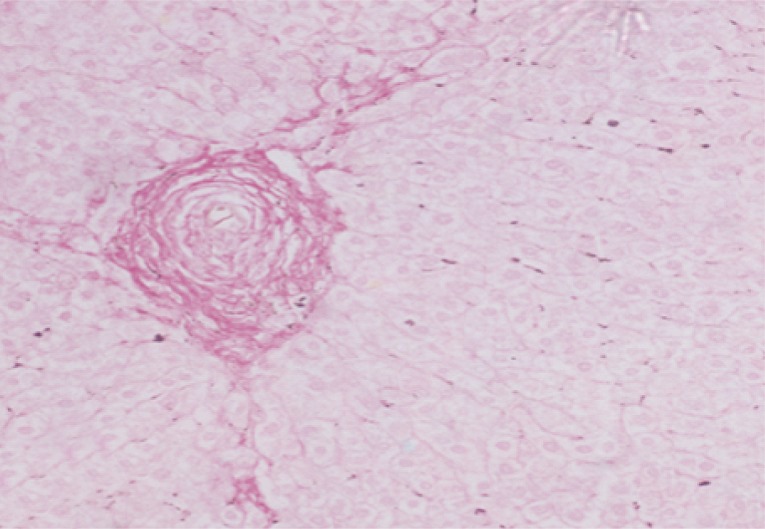
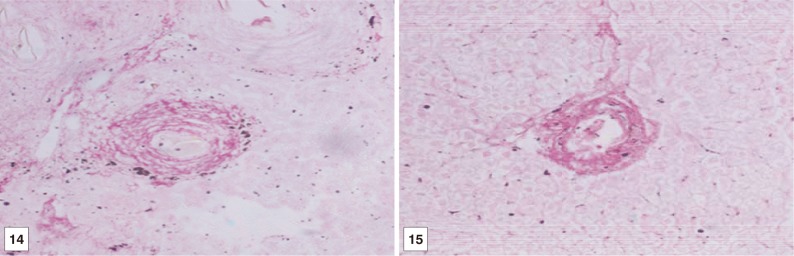
Granuloma count was carried out at a low power magnification (×10) of serial tissue sections of more than 250 µm a part in all fields of liver sections. The mean number of granulomas was calculated for each mouse. The mean was calculated for each experimental group from the mean values of the individual mice. Granuloma structural configuration, including cellular components and associated histopathological changes was also studied [23].
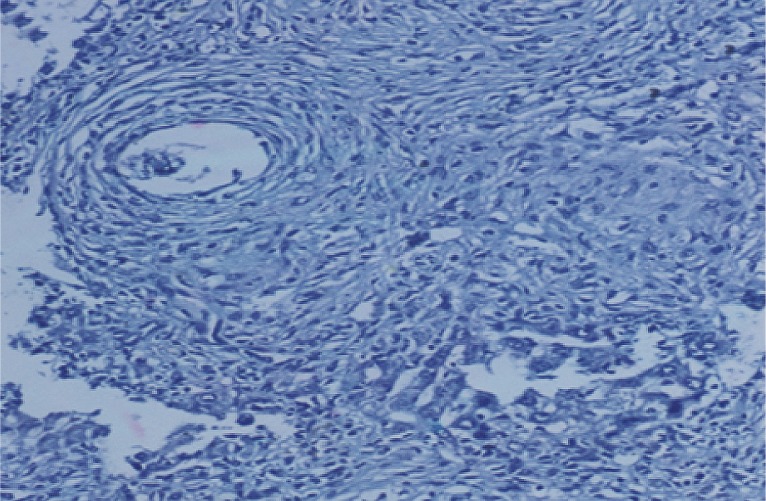
Sirius red staining for collagen
The collagen content of the liver sections stained with Picrosirius red was evaluated quantitatively for the percentage areas of collagen using Computerized Cell Image Analysis. Collagen fibers within the squares of the ocular reticule were counted at ×200 magnification [24].
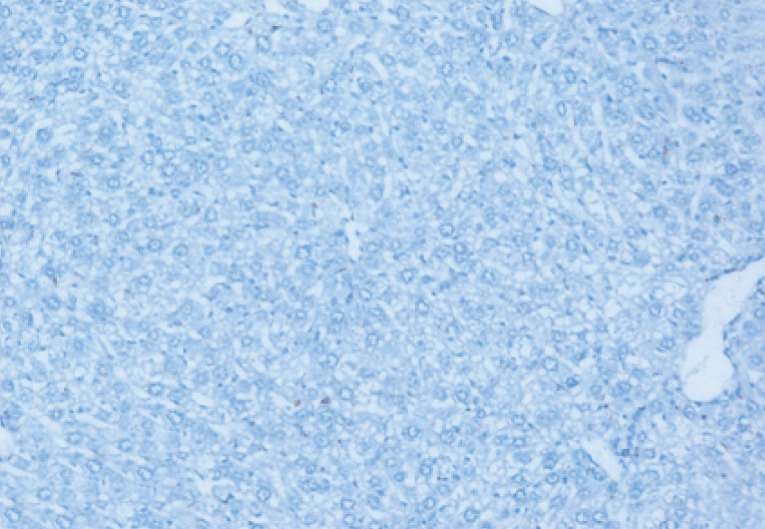
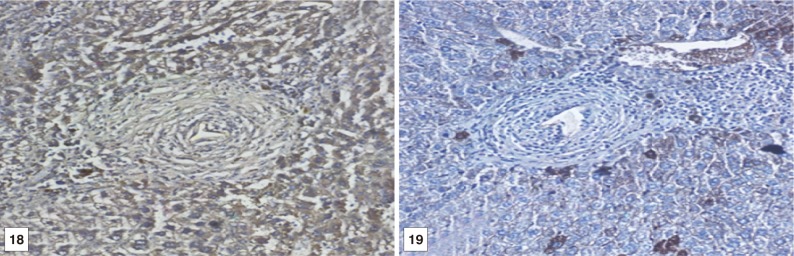
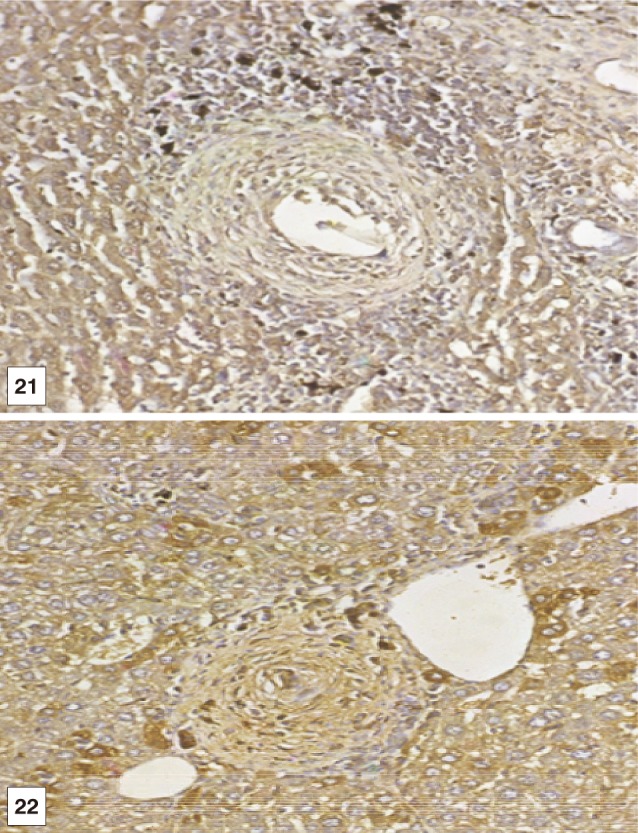
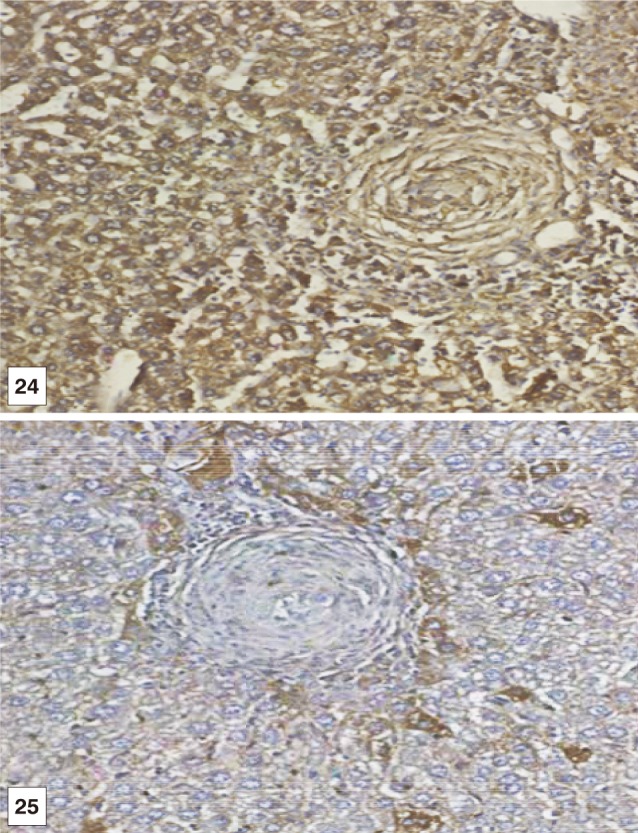
Detection of OV-6 in hepatic tissues using immunohistochemical technique
The presence of transplanted BMSCs in the liver tissue was analyzed by staining OV cells using a biotinylated anti-OV-6 antibodies followed by universal diaminobenzidine tetrahydrochloride (DAB) immunostaining detection kit. The standard avidin-biotin immunoperoxidase technique was used [25].
Liver function parameters
Serum albumin concentration: Formation of albumin/bromcresol-green complex at pH 4.2 and photometric measurement of the absorbance were detected by the Biodiagnostic kits [26].
Aminotransferase enzymes (ALT, AST): The alanine aminotransferase (ALT) and aspartate aminotransferase (AST) activities were determined in serum using the Biodiagnostic kit [27].
Alkaline phosphatase (ALP) activity: ALP was determined in serum according to the method described by Belfield and Goldberg [28], using the Biodiagnostic test kit.
Statistical analysis
The statistical analysis was performed using the SAS, ver.9.2 software program. Data were summarized as mean±standard error (SE) for values and percentages. For comparison of more than 2 groups, the analysis of variance, ANOVA test, was used. Comparison between percentages was calculated by the chi-square test. Significant differences between means were determined by Duncan Post Hoc test [29]. P<0.05 implies a statistically significant difference.
RESULTS
PCR detection of male-derived BMSCs
The sry gene that was used as the Y chromosome marker was detected in the liver tissue from the Schistosoma-BMSCs groups and Schistosoma-BMSCs-HGF groups, but not in the infected control or uninfected negative control groups (Fig. 1).
Liver function parameters
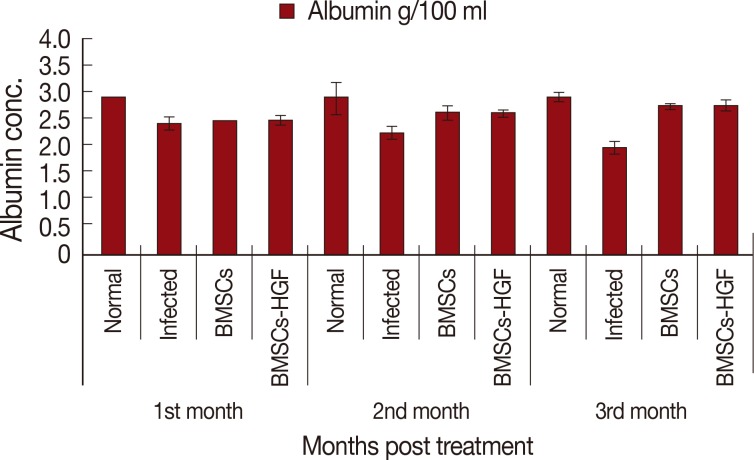
The performance of the liver was monitored by the serum level of liver proteins and enzymes as presented in Table 2 and Figs. 4, 5. At the 1st month PT, both groups treated with BMSCs only and BMSCs supplemented with HGF showed non-significant increases in serum albumin concentration as compared to the infected control group. At the 2nd and 3rd month PT, the albumin concentration in sera of mice treated with BMSCs and BMSCs supplemented with HGF increased significantly (P<0.05) compared to the infected control group.
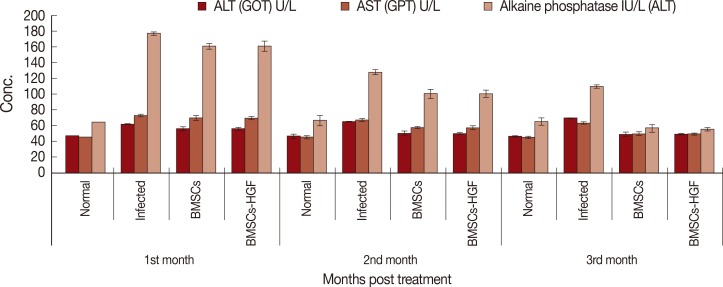
At the 1st month PT with both BMSCs only and BMSCs supplemented with HGF, there were significant reductions (P<0.05) in both ALT and ALP activities, whereas, this reduction was non-significant in serum AST activity as compared to the infected control group. At the 2nd and 3rd month PT, the activities of ALT, ALP, and AST in sera of mice treated with BMSCs and BMSCs supplemented with HGF was reduced significantly (P<0.05) in comparison with that of the infected control group.
Detection of OV-6 in hepatic tissues using immunohistochemical technique
The presence of OV-6 in liver sections of normal, S. mansoni-infected mice, and infected mice treated with BMSCs and BMSCs supplemented with HGF was detected immunohistochemically using monoclonal anti-mouse OV-6 antibodies as a marker for newly formed hepatocytes differentiated from BMSCs. Normal uninfected negative control and S. mansoni-infected control mice showed negative expression of OV-6 in hepatocytes and granuloma cells.
At treatment with BMSCs only and BMSCs in combination with HGF, positively stained hepatocytes with brown colour appeared indicating the presence of newly formed hepatocytes. As the period after treatment with BMSCs increased, the expression of OV-6 in hepatocytes also increased. The greatest increase in the expression of OV-6 was recorded in hepatocytes of mice treated with BMSCS at the 3rd month PT. The staining of cells occurred intercalated between mouse hepatocytes and around granulomas present either singly or in clusters (Figs. 16,17,18,19,20,21,22,23,24,25).
DISCUSSION
Schistosomiasis is a wide-spread chronic helminthic infection which contributes to death of over half a million people yearly. The Schistosoma-induced liver disease associated with granulomatous inflammation constitutes the most prevalent disease among Egyptians. The disease is characterized by disorganized proliferation of hepatocytes, excessive scarring, and excessive collagen deposition which lead to chronic liver failure. Hence, chronic schistosomiasis was selected as an experimental model of a chronic liver disease which allows evaluation of stem cell migration to sites of damage and their subsequent contribution to repair and host survival [8].
This study was intended to demonstrate the fate of murine BMSCs after inoculation into murine diseased liver, in an attempt to set an experimental model of in vivo hepatocyte differentiation from BMSCs.
Previous studies have revealed that the intensity of schistosomal infection is related to the degree of liver fibrosis and granulomatous reaction [30]. The number of granulomas was correlated with the severity of the disease. As granulomas develop in response to egg deposition in the tissue, it can be possible to assess the amount of eggs retained in the liver and intestine [8]. This is in agreement with the present histopathological finding which was confirmed by the increased number and diameter of granuloma, number of liver and intestinal eggs, and extensive accumulation of fibrous tissues.
After treatment with BMSCs alone or in combination with HGF, there was an improvement in the histopathological picture of the liver which included diminution in the number and diameter of granulomas and reduction in the fibrotic content. Similar to the present findings, Abdel Aziz et al. [13] focused on the antifibrotic effect of mesenchymal stem cells (MSCs) on the liver of S. mansoni-infected mice. In their model, once granulomas were formed around the S. mansoni ova, the rate of spontaneous change of the collagen content of the granuloma was low, thereby providing a relatively stable in vivo model for analyzing collagen turnover. The histopathological examination of the liver of the S. mansoni-infected mice showed characteristic granulomas, which were cellular and contained thick collagen bands. After MSCs infusion, the granulomas decreased in size and number and became less cellular, although the concentric fibrous bands were still discernible.
In our study, the extent of fibrosis was estimated by the quantitative morphometric analysis of the collagen content in Sirius red stained liver sections. The results revealed significant decreases in the collagen content after treatment with BMSCs alone or in combination with HGF. In this regard, Ghanem et al. [8] demonstrated that fibrosis developed during the chronic phase of granulomatous inflammation in murine schistosomiasis and it represented a protective function during infection by neutralization and sequencing egg antigens that can potentially damage host tissues. The inhibitory effect of inoculated BMSCs on the collagen deposition may be related to an enhancement of fibrotic degradation rather than a decrease in fibrosis synthesis.
Pretreatment with MSCs [30] significantly reduced liver fibrosis compared to untreated cells. Antifibrotic effects of MSCs have been previously documented by many authors [31,32], and this might have a contributing factor for the observed decline in the liver fibrosis. A significant reduction of collagen content [13] estimated by Sirius red staining in MSCs infusion group was recorded as compared to the PI control values. They [13] also reported that resorption of fibrous tissues was observed from the first month post BMSCs administration. This was more rapid than fibrous tissue resorption observed with anti-Schistosoma drug treatment, where resorption started 3 months after the treatment.
Oval cells are hepatic precursors of hepatocytes and bile duct cells. The OV cells originated from BMSCs [33]. Chronic viral hepatitis was investigated [34], and it was noted that similar hepatic OV cells were allocated predominantly in the periportal region and fibrosis septa, characterized by an ovoid nucleus, small size, and scanty cytoplasm. In a study carried out to investigate the potential of BMSCs in regeneration of hepatic lesions induced by S. mansoni chronic infection [1], it was reported that transplanted cells migrated to granuloma areas and the cytokine level associated with fibrosis deposition in liver fragments of mice submitted to therapy was reduced. They also reported that infection with S. mansoni caused an increase in the number of OV cells which were mainly found in periportal areas of the hepatic lobe in saline-treated as well as BMSCs-treated mice.
The results of the current study showed that OV cells constituted the dominant forms of newly formed cells observed in the grids of the treated group. Stem cells in this study had plenty of time to develop and differentiate as the experiment last for 3 moths post transfection with BMSCs. On the contrary, another author reported that inoculated stem cells lasted for hours after the acute liver injury [35,36].
In our study, there was an increase in OV cell number in liver sections of different study groups. Using the anti-mouse OV-6 marker, the liver sections of treated groups either with BMSCs or with BMSCs supplemented with HGF showed positive staining results with increase in the number of cells. Negative reaction for OV-6 marker was noticed in the liver sections of either normal uninfected negative control group or infected untreated groups.
S. mansoni infection resulted in a hepatocellular injury, which in turn led to a release of enzymes from the injured hepatic cells into blood circulation [37]. In this regard, serum biochemical profiles in terms of ALT, AST, ALP, and albumin provided supportive evidence to the pathological alterations observed in response to schistosomal infection.
Aminotransferases are considered as marker enzymes for cell toxicity and their elevated level in serum give an additional support for S. mansoni cytotoxicity. Consistent with previous studies, the present study showed that the inflammatory reactions induced in the livers of S. mansoni-infected mice were ensured by liver dysfunctions reflected by elevated serum levels of ALT, AST, and ALP. These high serum concentrations of ALT and AST were attributed to the hepatocellular damage resulting from egg deposition where the transaminase levels showed an intimate relationship with cell necrosis and/or increased cell membrane permeability to discharge of the enzyme to the blood stream [38].
Many authors attributed the elevated level of ALP to irritation of liver cells by toxins or metabolites from growing schistosomules and eggs or due to a loss of intracellular enzymes by diffusion through the cell membrane which appeared to act as a stimulus to the synthesis of more enzymes, which would in turn increase the rate of diffusion and hence increase the serum enzyme activity [39,40].
In the present study, supplementation of BMSCs effectively ameliorated the above serum marker levels characteristic of schistosomal hepatopathy, and the best results were observed at the 3rd month PT. S. mansoni infection causes hypoalbuminemia in mice, and this phenomenon occurs simultaneously with an increase in collagen deposition and could be associated with a decrease in albumin mRNA [41]. In the present study, albumin was clearly recognized in serum at the 3rd month PT, but the rate of collagen deposition in the tissue was not changed considerably. Therefore, it is recommended to follow up the results for longer periods PT.
Also, the present results were in accordance with Abdel-Rahim et al. [42] who reported that the low serum albumin level reflected an impaired protein synthesis due to liver destruction of parenchymal liver cells. The same results were also reported by Mahmoud et al. [43] who found a significant reduction in the albumin level due to S. mansoni infection.
After treatment with BMSCs, or BMSCs-HGF, the present results revealed significant increases in the serum albumin level at the 2nd and 3rd month PT in both groups as compared to the infected control group reaching normalization at the 3rd month PT. The autologous transplanted BMSCs may differentiate into hepatocytes, restore serum albumin, and suppress transaminase activity and liver fibrosis in the present experimental model of liver injury. There were borderline significant improvements in the serum albumin level of the transplanted group compared to the control group at the end of the 6th month PT [12].
A significant reduction in the serum ALP level was observed [44] in experimental animals transplanted with pretreated MSCs compared to the controls. The level of serum ALP has been used previously as an indicator of improved liver functions.
The increase in albumin and decrease in ALT and AST levels in the serum are indicative of normalization of the liver function by MSCs treatment [45]. The reversal of the functional parameters of the fibrotic liver could be either due to prevention of apoptosis of parenchymal cells or inhibition of proliferation and infiltration of inflammatory cells.
In agreement with the present results, BM-MSCs infused intravenously were localized to the liver and resulted in improvement of liver functions [13]. The improvement of liver functions has been observed with or without prior expansion, fractionation, and cloning of the MSCs or their differentiation into hepatocyte-like cells in culture. On the other hand, 4 weeks after BMSCs transplantation to mice with liver fibrosis induced by CCl4 administration, the mice showed significant reduction in liver fibrosis as assessed by improvement of liver functions, such as albumin and bilirubin levels and prothrombin time [46].
In conclusion, generally BMSCs with or without HGF infusion significantly enhanced hepatic regeneration and alleviated hepatic fibrosis in a S. mansoni-induced fibrotic liver model and has therapeutic effects which appeared in reduction of immunopathology, granuloma formation, and hepatic fibrosis related to S. mansoni infection. Therefore, it is important to use different types of stem cell factors other than HGF to show if there is a considered difference in the liver regeneration intensity. This infusion could generate new hepatocytes to improve the function of residual hepatocytes. Therefore, autologous BMSCs transplantation may be considered as a curative or supportive therapy for liver disorders.