Trichomonas vaginalis, a parasitic protozoan, causes the sexually transmitted disease trichomoniasis, a common infection of the urogenital system [1]. The World Health Organization (WHO) estimated that over a half of the 276.4 million new T. vaginalis infections occur each year in men [2]. Because trichomoniasis is asymptomatic in over 75% of infected men, it can remain undiagnosed and untreated, which has been hypothesized to result in chronic persistent prostatic infection [3,4]. In addition, T. vaginalis has been detected recently in the urine of patients with prostatitis and in the prostate tissue of patients with benign prostatic hyperplasia or prostate cancer [5–7]. However, the host-parasite interaction of trichomoniasis remains incompletely understood in the prostate.
Adhesion of the organisms to mucosal cells is considered a first and prerequisite step for T. vaginalis infections [8,9]. When T. vaginalis was added to epithelial cells, the parasitic body changed to a more elongated shape with pseudopodia, showing high adherence to primary vaginal epithelial cells. In contrast, the morphology of T. vaginalis did not change when added to a cervical cancer epithelial cell line (HeLa) [10]. Therefore, T. vaginalis is able to recognize and specifically bind to host tissues. Adhesion of T. vaginalis to different cell types such as renal epithelial cells, colon epithelial cells, sperm, fibroblasts, leukocytes, or breast myocytes has been previously reported [11–15]. However, adhesion of T. vaginalis to prostate epithelial cells has not yet been observed, although there are several reports on T. vaginalis-induced inflammation of prostate epithelial cells.
The aim of this study was to investigate whether adhesion and cytotoxicity of T. vaginalis could lead to inflammation of prostate epithelial cells. Our findings suggested that T. vaginalis adhered and caused cytotoxicity to prostate epithelial cells, with subsequent induction of inflammation and epithelial-mesenchymal transition (EMT).
T. vaginalis isolate T016 was grown in TYM media supplemented with 10% heat-inactivated horse serum. The human prostate epithelial cells (RWPE-1 cell line) were cultured in keratinocyte serum-free medium (K-SFM) containing 5 ng/ml human recombinant epidermal growth factor and 25 μg bovine pituitary extract supplemented with 5% FBS.
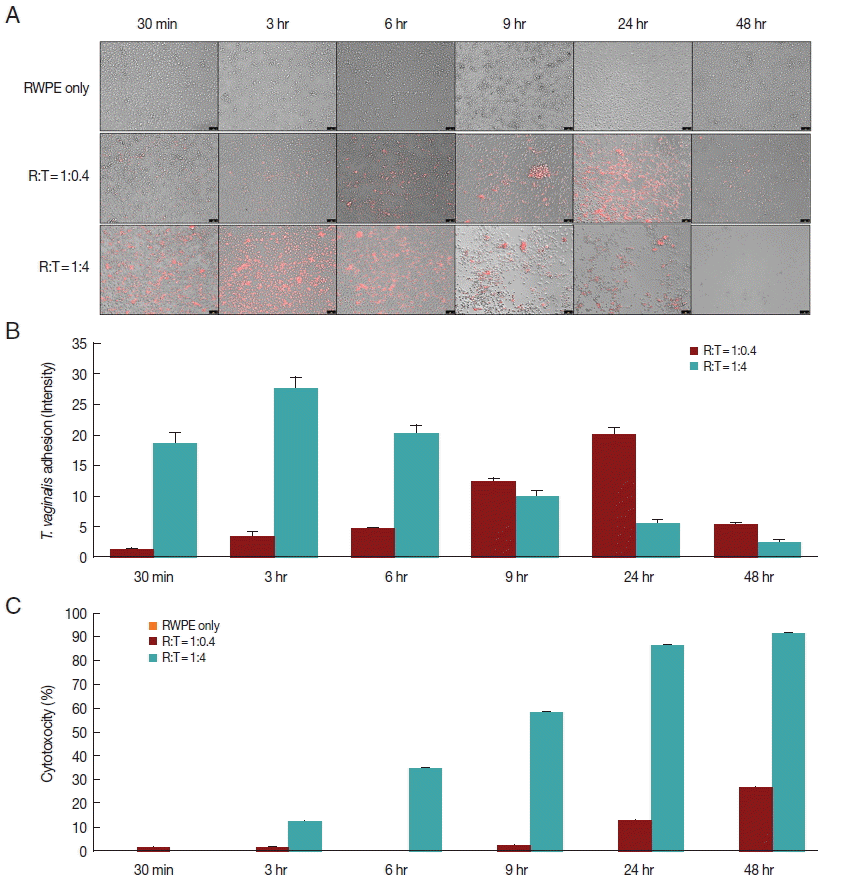
To confirm the adhesion of live T. vaginalis, RWPE-1 cells were incubated with T. vaginalis (RWPE-1: T. vaginalis=1:0.4 or 1:4) for 30 min, 3, 6, 9, 24, and 48 hr. T. vaginalis was stained with a CellTrackerOrange Fluorescent Probe (Invitrogen, Carlsbad, California, USA) according to the manufacturer’s instructions. Briefly, T. vaginalis was suspended in CellTrackerOrange solution, which was diluted to 0.5 μM in Hank’s solution (Invitrogen). After 30 min, the stained T. vaginalis was added to a monolayer of RWPE-1. After incubation, adherence of T. vaginalis was examined with a fluorescence microscope (Leica, Wetzlar, Germany), and images were analyzed by ImageJ.
To determine the effects of live T. vaginalis on the cytotoxicity of prostate epithelial cells, RWPE-1 cells were seeded on 96-well plates and cultured in complete K-SFM medium with 5% FBS. After 24 hr, the RWPE-1 medium was changed to serum-free RPMI1640 medium and incubated for 24 hr. T. vaginalis (RWPE-1:T. vaginalis=1:0.4 or 1:4) was then added into the RWPE-1 and incubated for various times. Cytotoxicity was analyzed using a CCK-8 assay. CCK-8 reagent (Enzo Life Sciences, Farmingdale, New York, USA) was added to each well for 2 hr prior to obtaining the spectrophotometric reading according to the manufacturer’s directions. The CCK-8 assay is a sensitive colorimetric assay to determine the number of viable cells in cell proliferation and cytotoxicity assays. The CCK-8 reagent is reduced by dehydrogenases, yielding a water-soluble formazan dye. The amount of dye generated is directly proportional to the number of living cells.
The production of inflammatory cytokines by RWPE-1 cells incubated with T. vaginalis (RWPE-1: T. vaginalis=1:0.4 or 1:4) for 9, 24, and 48 hr was measured using an ELISA for IL-1β, IL-6, CCL2, and CXCL8 (BD Biosciences, San Diego, California, CA). The data are presented as the mean±SD of 3 independent experiments.
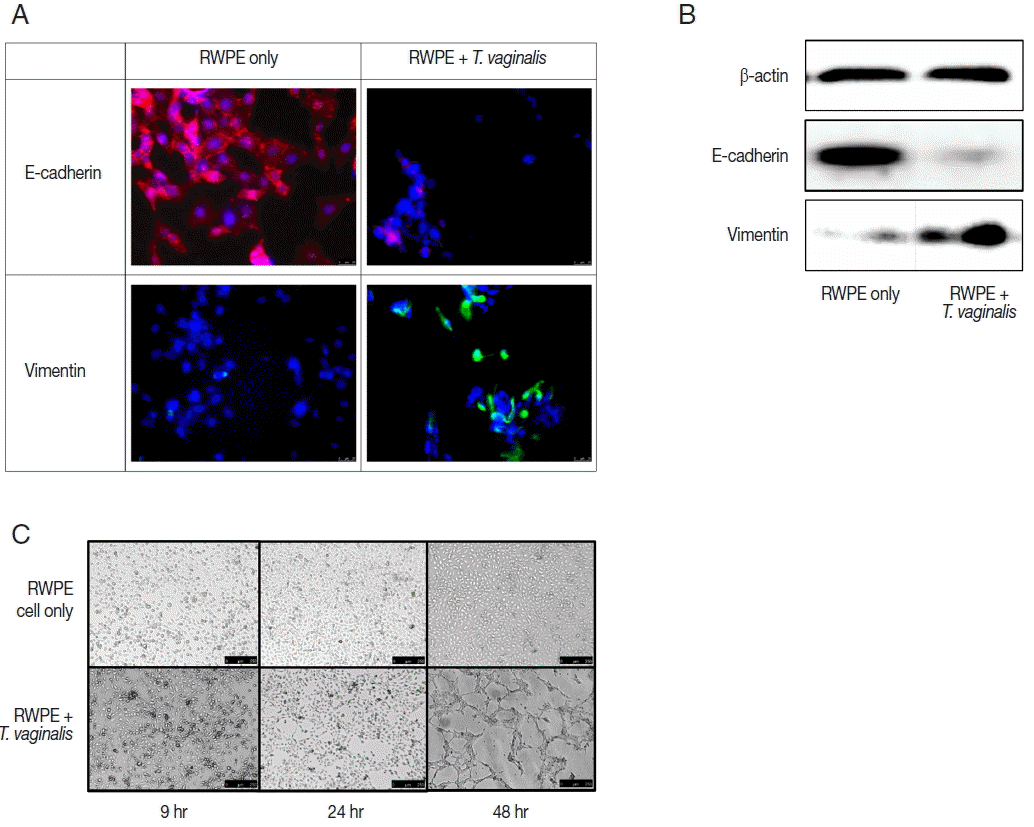
To examine the epithelial-mesenchymal transition (EMT), expression of E-cadherin (epithelial marker), and vimentin (mesenchymal marker) was assessed using fluorescence microscopy. Briefly, RWPE-1 cells were grown on a glass coverslip in a 12-well plate (Corning) and incubated with T. vaginalis for 24 hr. The cells were fixed with 4% paraformaldehyde at −20°C for 10 min and blocked with 0.1% normal goat serum at room temperature for 1 hr. After being washed with PBS 3 times, the glass cover slips were incubated with anti-E-cadherin antibody (1:250, mouse monoclonal, Cell signaling) or anti-vimentin antibody (1:500, rabbit polyclonal, Abcam) overnight at 4°C. They were then washed and stained with CyTM3 AffiniPure donkey anti-mouse IgG (1:500, Jackson Immuno Research, West Grove, Philadelphia, USA) or Alexa 594-labelled goat anti-rabbit lgG (#A11012, 1:500, Invitrogen) at 37°C for 1 hr. The cover slips were mounted using Vectashield mounting medium (Vector Laboratories, Burlingame, California, USA) and stained with DAPI to visualize the nuclei. Fluorescence was then measured with a fluorescence microscope (Las Software, Leica).
Western blotting was also performed to confirm EMT. RWPE-1 cells were incubated with T. vaginalis (RWPE-1: T. vaginalis= 1:0.4) for 48 hr, separated by SDS–PAGE on a 12% polyacrylamide gel, and transferred to an Immun-Blot® PVDF membrane (Bio-Rad, Quarry Bay, Hong Kong). The membranes were probed with anti-E-cadherin antibody (1:1,000; Cell signaling) or anti-vimentin antibody (1:1,000; cell signaling) primary antibodies overnight at 4°C. The membranes were incubated with goat anti-rabbit IgG polyclonal antibody (1:10,000; ADISAB-300-J, Enzo Life Sciences) for 1 hr at room temperature. The blots were visualized using Chemiluminescent Sensitive Plus HRP Microwell and/or Membrane Substrate (SurModics, Minneapolis, Minneapolis, USA). The signals were measured with a Chemi-Doc (Bio-Rad, Hercules, California, USA).
When prostate epithelial cells were reacted with T. vaginalis at a ratio of 1:0.4, adhesion of T. vaginalis began after 30 min and increased continuously for 24 hr. In contrast, at a ratio of 1:4, adherence of T. vaginalis to prostate epithelial cells peaked at 3 hr, after which the level sharply decreased (Fig. 1A, B). When RWPE-1 cells were infected with T. vaginalis (RWPE-1: T. vaginalis=1:0.4), the cytotoxicity of prostate epithelial cells increased gradually from 9 hr (2.0%) to 48 hr (26.4%). However, at a ratio of 1:4, the cytotoxicity of prostate epithelial cells increased from 3 hr to 48 hr, with a maximum level of 91.5% (Fig. 1C). In contrast to T. vaginalis-infected groups, cytotoxicity did not appear over a 48 hr period in the control group (RWPE only). This result was consistent with the results of Lustig et al. [16], who identified T. vaginalis contact-dependent cytolysis of epithelial cells. The 1:0.4 ratio showed higher adhesion and lower cytotoxicity at 24 hr. T. vaginalis is a microaerophilic anaerobic protozoa [17]. In contrast to the vagina, the prostate is assumed to be an anaerobic condition. In addition, high concentration of zinc in the prostate may induce a critical cytotoxicity to T. vaginal [18]. Considering that prostate tissue is a harsh condition for T. vaginalis survival, only a few T. vaginalis organisms remained viable in the tissue. Thus, 1:0.4 was chosen to determine cytokine levels and EMT.
To investigate the production of inflammatory cytokines by infected prostate epithelial cells, RWPE-1 cells were incubated with T. vaginalis (1:0.4) for 6, 24, and 48 hr. The RWPE-1 cells released the cytokines IL-1β, IL-6, CCL2, and CXCL8. Cytokine production was significantly greater in the infected group compared to the control group (Fig. 2). Production of IL-1β and CCL2 increased in a time-dependent manner (Fig. 2A, C), while the production of IL-6 and CXCL8 increased for 24 hr and subsequently decreased (Fig. 2B, D).
Several researchers have reported that the adhesion of T. vaginalis to vaginal epithelial cells plays an important role in the pathogenesis of trichomoniasis through production of CXCL8, CCL2, and IL-6 [9,19,20]. In previous studies, we reported that T. vaginalis-infected prostate epithelial cells produced pro-inflammatory cytokines such as IL-1β, IL-6, CCL2, and CXCL8 [21,22], which is consistent with the present results (Fig. 2). Therefore, these pro-inflammatory cytokines may be produced as a result of adhesion of T. vaginalis to prostate epithelial cells, which could promote the pathogenesis of prostatic disease. However, further study is required to determine whether inhibition of adhesion with anti-adhesin antibody or chemicals affect cytotoxicity and cytokine production. Also, considering invasiveness of T. vaginalis, it is interesting to examine whether adhesion of trichomonads to prostate stromal cells might be related to stromal inflammation or proliferation.
Expression of EMT markers was measured by fluorescence microscopy and western blot. In this study, we confirmed EMT through decreased expression of E-cadherin (an epithelial cell marker) and increased expression of vimentin (a mesenchymal cell marker) (Fig. 3A, B). EMT is a biological process that allows a polarized epithelial cell to undergo multiple biochemical changes enabling it to assume a mesenchymal cell phenotype. Enhanced migratory capacity, invasiveness, elevated resistance to apoptosis, and greatly increased production of extracellular matrix components occur in EMT [23]. EMT may be classified into 3 subtypes. Type 1 EMT involves the transition of primordial epithelial cells into motile mesenchymal cells and is associated with the generation of diverse cell types during embryonic development and organogenesis. Type 3 EMT occurs in carcinoma cells. Type 2 EMT involves transition of secondary epithelial cells to resident tissue fibroblasts and is associated with wound healing, tissue regeneration, and organ fibrosis [23–25]. Also, type 2 EMT is induced in response to inflammation, but stops once inflammation is attenuated, especially during wound healing and tissue regeneration [24]. In the present study, EMT of prostate epithelial cells was induced by T. vaginalis at 24 and 48 hr, along with increased cytotoxicity and cytokine production. Thus, our results indicate that EMT may be classified as type 2, which suggests a healing process in wounded prostate epithelial cells.
The ability of epithelial cells to change morphology to more closely resembled mesenchymal cells is a feature of EMT. Tube formation in vascular endothelial cells observed in cancer tissues is indicative of metastasis [26]. Maniotis et al. [27] showed that vascular mimicry patterns form in vitro in the absence of hemodynamic forces, suggesting that the formation of a patterned non-endothelial cell-lined microcirculation may be an attribute of the tumor cell. In this study, tube-like formation was observed after 48 hr of incubation (Fig. 3C). Morphologic changes of prostate epithelial cells induced by EMT may lead to tube-like formation. We showed previously that T. vaginalis induced EMT in prostate epithelial cells [22]. In this study, we confirmed additional morphologic changes, as well as EMT.
In conclusion, adherence of T. vaginalis to prostate epithelial cells caused cytotoxicity, pro-inflammatory cytokine production, and EMT. Our findings suggested for the first time that T. vaginalis may induce inflammation via adhesion to normal prostate epithelial cells.