Toxoplasma gondii is an obligate intracellular protozoan parasite with a global distribution in humans and other mammals. Although this parasite most often causes subclinical infection, primary infection during pregnancy can induce fetal damage and abortion in both humans and animals. T. gondii is a well-known inducer of type 1 cytokines, and while these cytokines are required to survive infection, their over-production can be harmful and even lethal [1,2].
Mast cells are abundant in the skin, mucosa of the digestive and respiratory tracts and conjunctiva. These cells provoke IgE-mediated allergic responses and are involved in the control of helminth infections of the gastrointestinal tract. They play a key role in innate and adaptive immunity [3,4] and have been shown to induce acute inflammatory reactions in mice in response to intraperitoneal injection of T. gondii [5]. Mast cells also protect the host when T. gondii is introduced by the oral route [3]. Mediators produced by activated mast cells may reduce acute inflammation and stimulate parasite clearance in T. gondii infections [6]. On the other hand, T. gondii itself inhibits mast cell degranulation by suppressing the mobilization of intracellular Ca2+ by phospholipase C [7]. So, the overall effect of mast cells on T. gondii infections is uncertain. In addition, it is not known whether T. gondii lysates can induce inflammatory responses in the mast cell leukaemia cell line, HMC-1. The aim of this study was to evaluate whether T. gondii lysates induce the activation of HMC-1 cells to release cytokines and nitric oxide.
T. gondii (RH strain) tachyzoites were maintained by intra-peritoneal infection of ICR mice (Osan, Korea). Tachyzoites were harvested from the mice, washed with PBS and centrifuged at low and high speeds (500 g and 3,000 g) for 5 min to remove peritoneal cells. To prepare T. gondii lysate, the tachyzoites were frozen and thawed 3 times and ultrasonicated at 100% amplitude, 0.75 cycles, 7–10 times, until the tachyzoites were completely disrupted. The sonicate was centrifuged at 10,000 g for 1 hr, and the supernatant was filtered through a 0.2 μm membrane (Millipore, Bedford, Massachusetts, USA). Protein concentrations were determined with the Bradford assay (Bio-Rad, Hercules, California, USA).
Human leukemic mast cells (HMC-1) were grown in IMDM supplemented with 10% FBS at 37°C in a 5%-CO2 incubator. To examine cytokine and iNOS mRNA expression, HMC-1 cells (1.5×106/well) were pretreated with IFN-γ (5 μg/ml, ProSpec, East Brunswick New Jersey, USA) for 1 hr and then stimulated with an aliquot of T. gondii lysate (50 μg/ml) or LPS (10 μg/ml, Sigma, St Louis, Missouri, USA). As a positive control, the HMC-1 cells were incubated with PMA (100 nm/ml) and A23187 (10 μm/ml). Total RNA was extracted from the cells using Trizol reagent (Invitrogen, Carlsbad, California, USA) and 200 μl chloroform (Sigma Aldrich, St. Louis, Missouri, USA). cDNAs were synthesized and reverse-transcribed using RT Premix and PCR Premix (iNtRON Biotechnology, Seongnam, Korea). Primer sequences were as follows; β-actin (5′-CCA GAG CAA GAG AGG TAT CC-3′ and 5′-CTG TGG TGG TGA AGC TGT AG-3′), human IL-8 (5′-GCC AAG AGA ATA TCC GAA CT-3′ and 5′-AAA GTG CAA CCA CAT GTC CT-3′), MCP-1 (5′-TCT GTG CCT GCT GCT CAT AG-3′ and 5′-GCT GCA GAT TCT TGG GTT GT-3′), IL-4 (5′-CCC TTT CGG CAA AAT CCT CC-3′ and 5′-AGG AAT ACA GTG CAG CTT ACC A-3′), TNF-α (5′-ACT CTT CTG CCT GCT GCA CTT TGG-3′ and 5′-GTT GAC CTT TGT CTG GTA GGA GAC GG-3′), iNOS (5′-AGC ATG AGC CCC TTC ATC AAT-3′ and 5′-CTG TTT CAA CGA CCT CCG GG-3′), and GAPDH (5′-GTC AGT GGT GGA CCT GAC CT-3′ and 5′-AGG GGT CTA CAT GGC AAC TG-3′). PCR conditions were initial DNA denaturation at 94°C for 5 min followed by 35 rounds of denaturation at 98°C for 15 sec, and annealing for 30s at 58°C for β-actin, GAPDH & iNOS, 56°C for IL-8 & IL-4, 55°C for MCP-1, and 62°C for TNF-α, and extension at 72°C for 35 sec.
To measure IL-8, MCP-1, TNF-α and IL-4 proteins, HMC-1 cells were pretreated with IFN-γ followed by exposure to T. gondii lysate or LPS for 18 hr or 24 hr. Supernatants were collected and stored at −20°C and cytokines were measured by ELISA (BD Biosciences, San Diego, California, USA).
Western blot analysis with monoclonal antibodies was used to detect expression of IL-8 receptor (CXCR1 and CXCR2) on HMC-1 cells which were pretreated with IFN-γ for 1 hr followed by T. gondii lysate or LPS. After incubation, the cells were harvested and lysed in PRO-PREP protein extraction solution (iNtRON Biotechnology, Seongnam, Korea). Equal amounts of protein were denatured and separated by SDS–PAGE on 10% polyacrylamide gels and transferred to Immune-Blot1 PVDF membranes (Bio-Rad, Quarry Bay, HongKong). The membranes were probed with anti-CXCR1 and anti-CXCR2 antibodies (1:1,000; Abcam, Cambridge, UK), or β-actin polyclonal antibody (1:3,000; Abcam, Cambridge, Massachusetts, USA) overnight at 4°C followed by goat anti-rabbit IgG polyclonal antibody (1:10,000; ADISAB-300-J, Enzo Life Sciences, New York, USA) for 1 hr at room temperature. The blots were visualized using Chemiluminescent Sensitive Plus HRP Microwell and/or Membrane Substrate (SurModics, Minneapolis, Minnesota, USA), and signals were measured with a Chemi-Doc1 (Bio-Rad).
For measuring nitric oxide, HMC-1 cells was pretreated with IFN-γ followed by T. gondii lysate or LPS for 3 hr, 6 hr, 18 hr or 24 hr and supernatants were collected. NO was determined as nitrite using Griess reagent [8].
Statistical analyses were performed with SPSS, version 21 (IBM, Chicago, Illinois, USA). The Mann-Whitney U test was used to compare results, and P-values<0.05 were considered statistically significant. Data are expressed as means±SDs of three independent experiments.
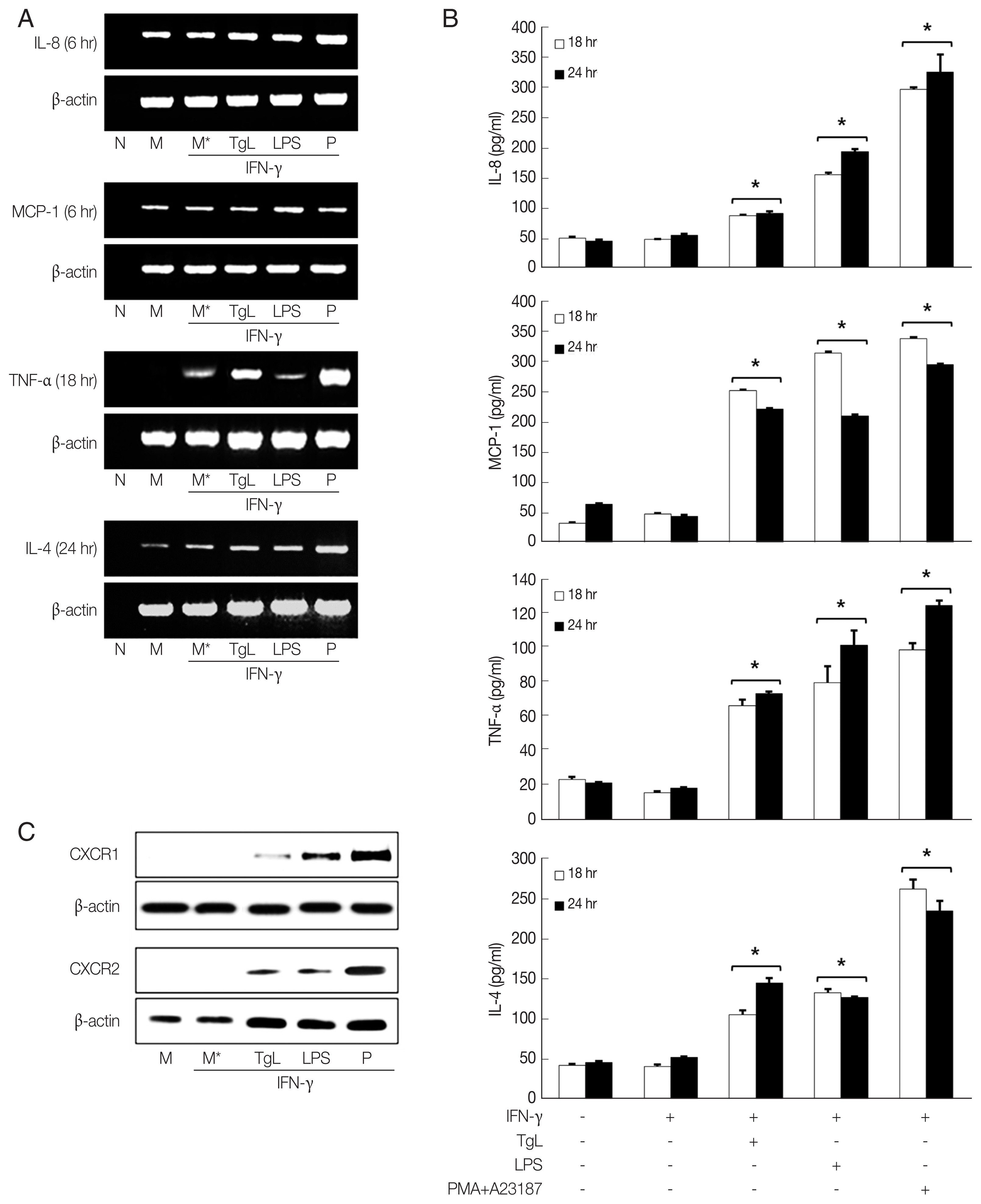
In this experiment, up-regulation of IL-8 mRNA and increased secretion of IL-8 were observed in the T. gondii-treated HMC-1 cells 6 hr and 18 hr post inoculation (PI), respectively. However the IL-8 levels of HMC-1 cells stimulated with LPS as positive control were significantly higher than those of the of T. gondii-stimulated cells. Up-regulations of MCP-1 mRNA and protein were also observed in the HMC-1 cells exposed to T. gondii lysate or LPS. Similarly, enhanced TNF-α mRNA expression and increased TNF-α secretion were noted in supernatants of HMC-1 cells treated with T. gondii lysate. IL-4 mRNA was also increased by incubation with T. gondii lysate for 24 hr, and IL-4 protein increased in the supernatants of treated HMC-1 cells (Fig. 1A, B). To investigate the expression of chemokine receptors for IL-8, CXCR1 and CXCR2 production was analyzed by western blotting after treatment of HMC-1 cells with T. gondii lysate or LPS for 1 hr. Both CXCR1 and CXCR2 increased in response to T. gondii lysate (Fig. 1C).
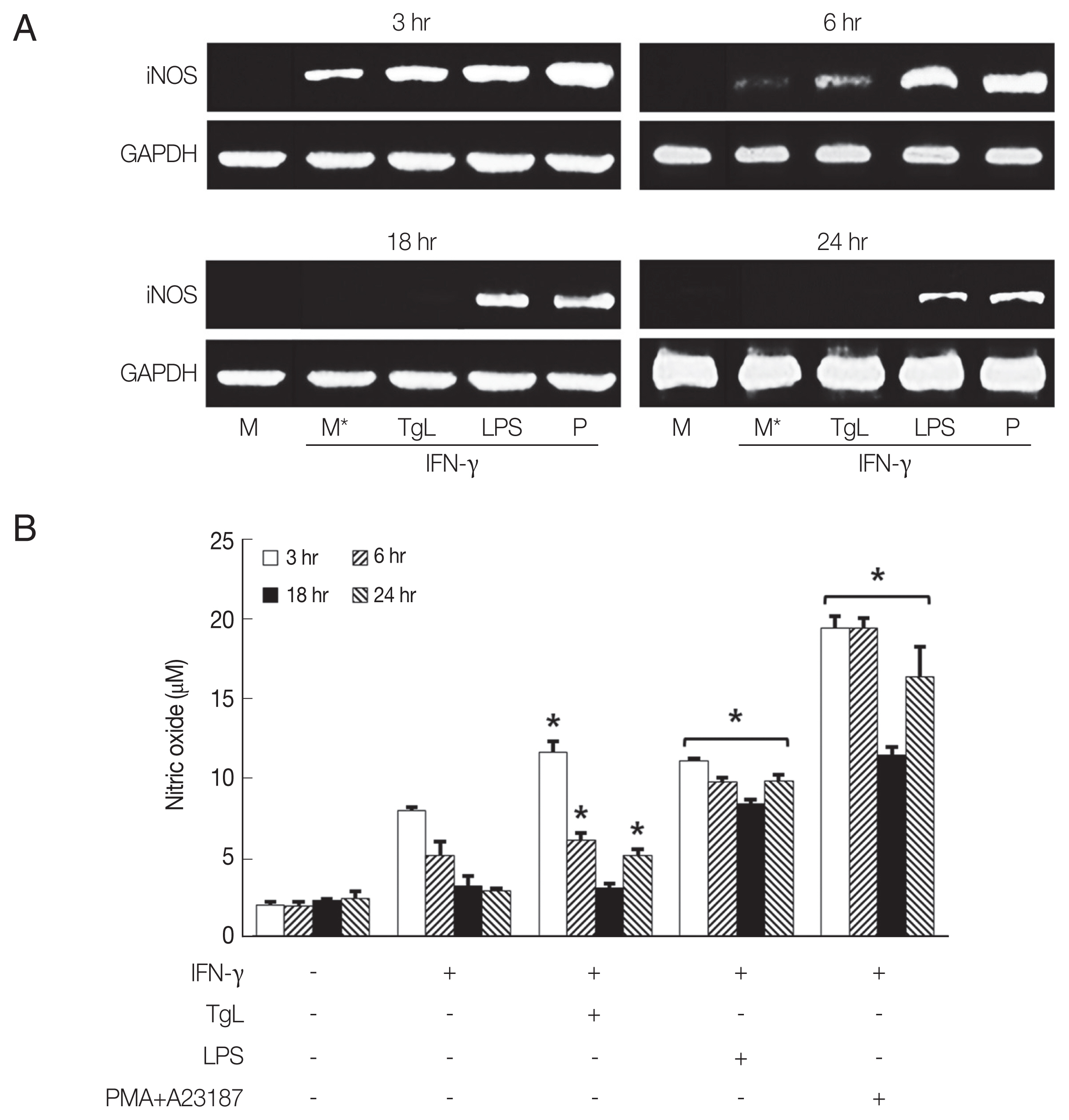
The results in Fig. 2A show that iNOS mRNA increased in HMC-1 cells treated with T. gondii lysate for 3 hr, and then gradually disappeared. In LPS-treated cells, iNOS mRNA continued to increase between 3 hr and 24 hr of incubation although at a decreasing rate. Maximal NO production in HMC-1 cells treated with T. gondii lysate or LPS was observed after 3 hr of stimulation (Fig. 2B).
In T. gondii infections, immune cells including neutrophils, monocytes and macrophages are activated, and also these cells are targets of T. gondii [9,10]. T. gondii down-regulates type I immune responses, blocking production of pro-inflammatory cytokine including IFN-γ in macrophages and dendritic cells [11]. Mast cells are abundant in the intestinal mucosa and play an important role in protective immune responses to oral infection by T. gondii [3]. Exposure to T. gondii causes mast cell to activate, degranulate and release mediators, proteases and cytokines [12]. Increases in mast cells numbers may be involved in the balance between inflammation and wound healing [13].
In the current study we found that IL-8 and MCP-1 secretion was up-regulated after 18 hr exposure to T. gondii lysate. This result is in agreement with our previous report that infection with T. gondii tachyzoites leads to increased secretion of IL-8 and MCP-1 by HeLa cells and fibroblasts [14]. Up-regulation of TNF-α and IL-4 mRNAs was observed in HMC-1 cells treated with T. gondii lysate for 18 hr and 24 hr, respectively. Production of both cytokines increased in T. gondii lysate- or LPS-treated HMC-1 cells indicating that T. gondii lysate induces inflammatory effect in the cells. T. gondii lysate alone did not increase cytokine production in HMC-1 cells. However, pre-treatment of IFN- γ resulted in increased cytokine production in the cells. Therefore, IFN-γ was thought to be acted as a trigger for HMC-1 cells in cytokine production by T. gondii lysate.
The functions of chemokines induced by intracellular pathogens include leukocyte recruitment, cell mediated immunity and anti-protozoal activity. IL-8 induces a rapid and transient increase of Ca2+ in mast cells. HMC-1 cells express the CXCL8 chemokine receptors, CXCR1 and CXCR2 [15–18]. Increased levels of these receptors were observed on HMC-1 cells stimulated with T. gondii lysate for 1 hr (Fig. 1C).
The release of mediators by mast cell activation plays an important role in modulating inflammation during T. gondii infection [6,16]. In protozoal infections these mast cells are characterized by degranulation and the production of pro-inflammatory mediators, such as TNF-α [13]. The mast cell activator, compound 48/80, up-regulated Th1 cytokine mRNA (IFN-γ, IL-12p40, TNF-α), and the mast cell stabilizer, disodium cromoglycate, increased Th2 cytokine mRNA (IL-4, IL-10) [12].
Mast cells are effector cells involved in clearance of parasites including Trichomonas vaginalis (T. vaginalis) [6]. We observed that rat peritoneal mast cells (RPMCs) were involved in the inflammatory response to T. vaginalis infection. Increased release of histamine and TNF-α were observed in RPMC-stimulated T. vaginalis ESP or live trichomonads [19]. Activation of mast cells has been reported to trigger production of cytokines and inflammatory mediators by protozoan parasites such as Plasmodium sp., Leishmania sp., and T. gondii [12].
Several protozoan parasites have evolved strategies to evade NO-mediated anti-microbicidal activity. Inducible nitric oxide synthase (iNOS)-derived NO can inhibit leucocyte adhesion in the micro-vasculature during ongoing inflammation. T. gondii infection partially inhibits NO production by murine macrophages and thus promotes parasite survival in phagocytic cells [12,20,21]. Leishimania sp. and Trypanosoma cruzi are known to inhibit iNOS expression [22,23]. We investigated HMC-1 cells were treated with T. gondii lysate to determine whether T. gondii infection affects NO production. Increased iNOS mRNA and NO production were observed at 3 hr PI and it then declined by 6 hr.
Mast cells induce recruitment of other immune cells to initiate inflammation. In the present study, we observed that T. gondii lysate stimulated the production of chemokines (CXCL8, MCP-1), cytokines (TNF-α, IL-4) and NO production in HMC-1 cells. Therefore, activated mast cells by T. gondii lysate may be involved in innate and adaptive immune responses against T. gondii infection.