Echinococcus granulosus sensu lato (s.l.) is a zoonotic parasite that infects a wide range of hosts, including sheep, dogs, cattle, and humans, and causes cystic echinococcosis (CE). To maintain their lifecycle, E. granulosus s.l. requires 2 mammalian hosts, the definitive host (dogs and other canids) and an intermediate host (sheep, cattle, camels, and goats) [1]. Echinococcosis is an important issue for farming industries due to its economic and public health impact [2]. Molecular taxonomic studies of E. granulosus s.l. have been conducted using mitochondrial DNA regions such as the cytochrome c oxidase subunit 1 (cox1) and NADH dehydrogenase subunit 1 (nad1) genes, which have been used to define the genotypes G1–G10 [3–6]. However, the features of G9 remain unclear [7,8]. These genotypes are currently used as standards for molecular classification and epidemiology analyses [9]. In addition, it has been recently shown that G2 is no longer a valid genotype as well, but micro variant of G3 is available [10].
In Central Asia, most of the agricultural lands are semi-arid, but there are mountain grasslands; therefore, the farming industries in the countries of this region are concentrated on pasture-based livestock production [11]. This provides favorable conditions for the transmission of Echinococcus species from livestock reservoirs. Since the collapse of the Soviet Union in 1991, CE has emerged as a major zoonosis in the region, and the incidence of CE in humans has significantly increased [11]. In Kazakhstan, studies of the prevalence of CE suggested that the Echinococcus sp., infection rates are 30–50% in sheep, 7% in cattle, 5–10% in strait dogs (final host), and >20% in shepherd dogs [12]. Most infected dogs have the G1 genotype, but the G6–G7 genotype has also been detected in dog feces. In Iran, genotypes G1–G3 are the dominant types responsible for most human and animal infections [13,14]; G6 is the second most prevalent genotype in the region, and it is increasingly being detected in human cases. In Uzbekistan, the prevalence of CE in sheep increased from 45% in 1990 to 62% in 2002 [11]. Although more than 8,000 people have been treated for cystic echinococcosis since 2002 in the 14 states of Uzbekistan [15], few, if any, epidemiological investigations have been conducted. Epidemiological investigations are key for establishing the epidemiologic relationships among circulating isolates. The information gathered should be considered in the development of local cystic echinococcosis control and prevention programs. Therefore, this information has significant epidemiological relevance because of the wide distribution of Echinococcus species.
The aim of this study was to investigate the dominant genotypes of E. granulosus s.l. isolates from various hosts in Uzbekistan. Genotypic and phylogenetic analyses of E. granulosus s.l. isolates obtained from human patients, a jackal, sheep, and a dog in Uzbekistan were conducted to determine their genetic relationships.
The fluid from 52 hydatid cysts (10 from lungs and 42 from livers) were collected from patients who underwent medical treatment at Samarqand Hospital in Uzbekistan in 2016–2017. Approval was obtained from the patients for the use of the hydatid cyst fluid for genetic analysis via ethic committee of ISAEV institute. The protoscoleces were obtained after centrifugation of the hydatid cyst fluid for 5 min at 2,000×g and were stored at −20°C until use. Many adult worms were obtained from the small intestine of a jackal by hunting at a nearby hill in Samarqand, Uzbekistan at the beginning of 2016. Six hydatid cysts were collected from the livers of sheep that were infected with E. granulosus s.l. in Samarqand, Uzbekistan in the winter of 2017. Many adult worms were also collected from a dog in Samarqand, Uzbekistan over the same time period. Each 10 adult worms from dog and jackal were stored −20°C until use. Total genomic DNA was extracted from all of the collected samples with the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The extracted genomic DNA was stored at −20°C until use. PCR was conducted using primer sets for cox1 and nad1 as described previously [16]. After purifying the PCR products with the ExpinTM PCR SV and ExpinTM Gel SV kits (GeneAll Biotech, Seoul, Korea), the cox1 and nad1 PCR products were analyzed by sequencing and phylogenetic analysis to confirm the genotype. The DNA sequencing was performed by Cosmo Genetech Co. (Cosmo Genetech, Seoul, Korea) using the 3730 xl DNA Analyzer (Applied Biosystems, Foster City, California, USA). The reference sequences of the cox1 and nad1 genes of E. granulosus s.l. genotypes G1–G10 (except G9) were obtained from GenBank (https://www.ncbi.nlm.nih.gov/nucgss). Isolate and reference sequences were used in a multiple alignment, and phylogenetic distances were calculated using MEGA version 6.0. Phylogenetic trees were constructed using the neighbor-joining method in MEGA version 6.0. The all sequences were uploaded to GenBank and assigned GenBank number as cox1; MK 975892-MK975951, nad1; MN696570-MN696629).
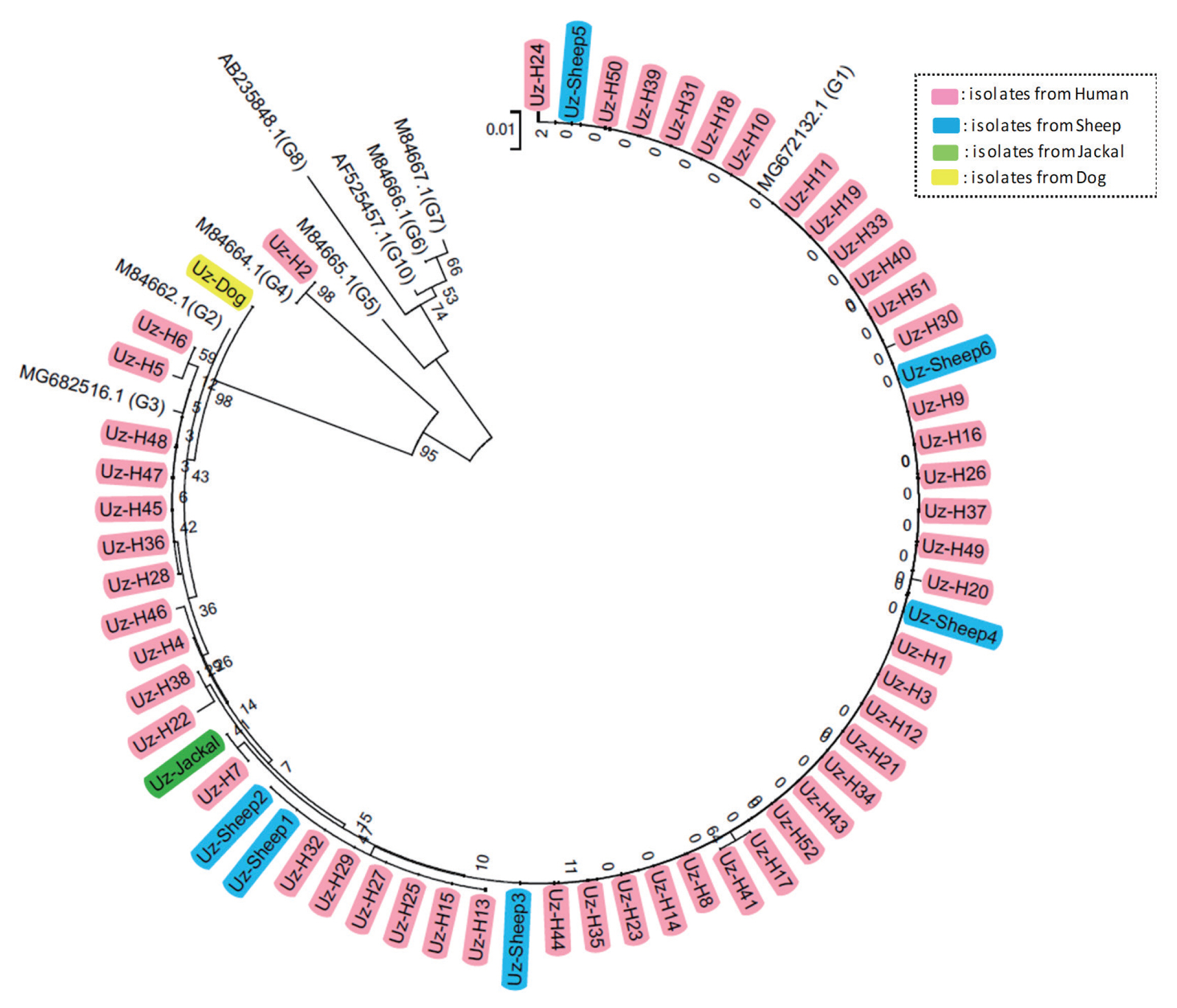
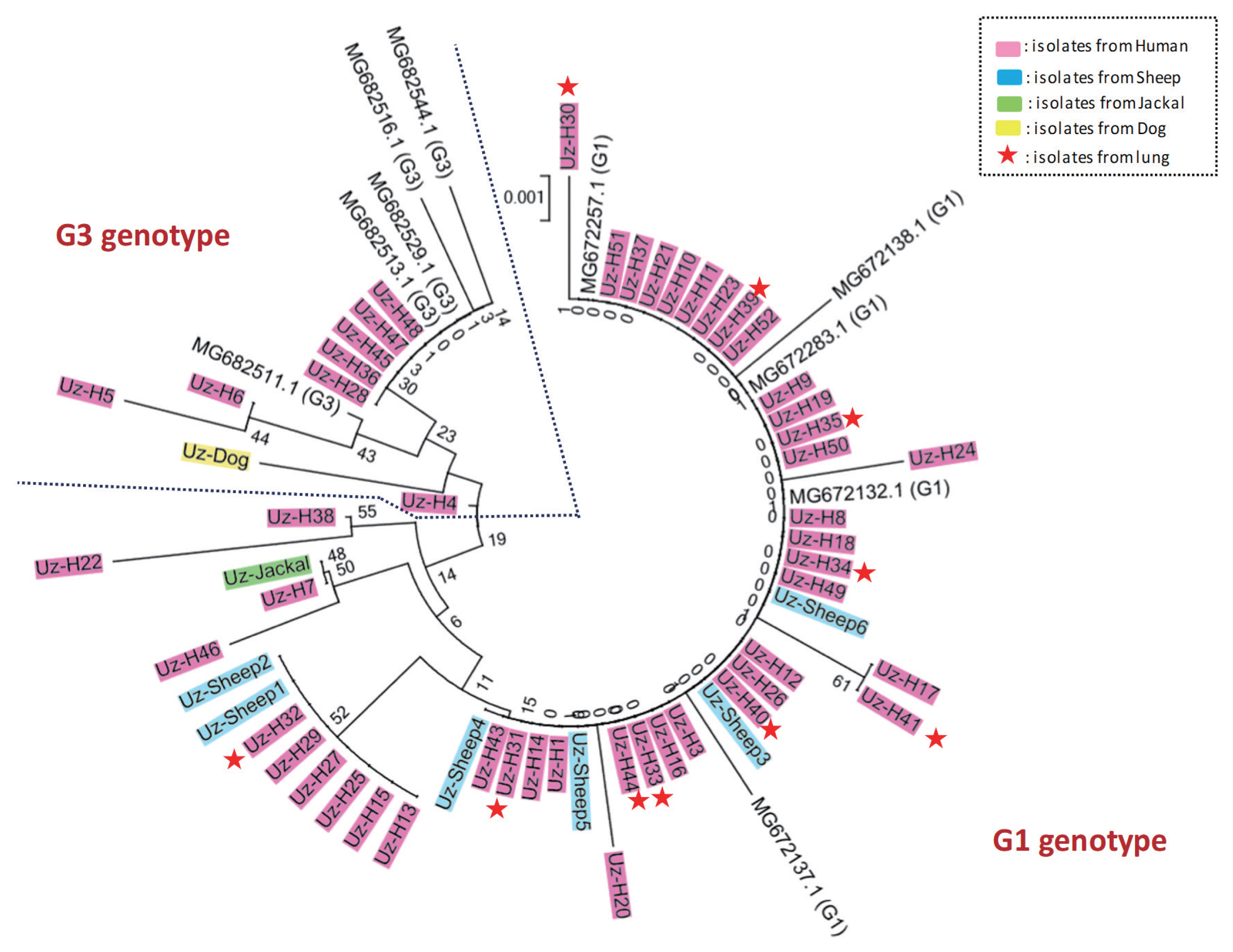
Various protoscolex samples were obtained from final and intermediated hosts in Uzbekistan, including 52 samples from human patients, 1 sample from a jackal, 6 samples from sheep, and 1 sample from a dog. To identify the genotypes of the E. granulosus in the samples, we performed sequencing and phylogenetic analysis. Sequence analysis of the cox1 and nad1 PCR products, including a multiple sequence alignment and phylogenetic analysis with reference sequences indicated that most of the isolates, except 1 sample (Uz-H2), had extremely high homology with genotype G1 and G3 E. granulosus s.s. reference sequences (Fig. 1, Supplementary Fig. S1). In contrast, Uz-H2 had extremely high homology with genotype G4 (E. equinus) reference sequences. However, the genetic distance between genotypes G1 and G3 is small, and thus, they could not be differentiated. To confirm the genotype (i.e., G1 or G3), we performed a detailed phylogenetic analysis using 5 G1 and 5 G3 isolates that were confirmed by near complete mitochondrial genome sequences in previous reports. As expected, 42 isolates from humans, all 6 isolates from sheep, and the 1 isolate from jackal were grouped with the confirmed G1 genotype sequences, while the remaining 8 isolates from humans and the 1 isolate from a dog were grouped with confirmed G3 genotype sequences (Fig. 2). To determine if the genotype of E. granulosus s.s. (i.e., G1 or G3) affects the organs that it can infect, we compared the genetic characteristics of the isolates according to the location of the cysts (10 from human lungs and 48 from the livers of humans and sheep). Interestingly, all isolates from the human lung grouped with confirmed G1 genotype strains.
Cases of CE in humans have been reported worldwide for more than 50 years. Until recently, the genus Echinococcus consisted of 9 valid species, including the livestock-friendly species [E. granulosus s.s. (G1 and G3), E. equinus (G4), E. ortleppi (G5), and E. canadensis (G6 and G7); species name is controversial ] and the wild animal-friendly species [E. canadensis (G8, G10), E. felidis, E. multilocularis, E. shiquicus, E. vogeli, and E. oligarthra] [7,8,17–19]. It is worth noting that genetic classification of isolates from both definitive and intermediate hosts can contribute to programs aimed to control this public health problem by determining how these organisms maintain their life cycle. Generally, there are many more genetic studies on livestock-friendly species because of their commonness. Previous studies revealed that E. granulosus s.s. G1 and G3 genotypes are the most common one infecting humans [20]. In fact, E. granulosus s.s. G1 and G3 genotypes are the most common worldwide (72.9%), followed by G6 and G7 genotypes (12.2% and 9.6%) [20]. In the Middle East, including in Iran and Kazakhstan, the G1 genotype is detected at a high frequency [12,21–23]. In this study, we firstly analyzed the genetic variation of E. granulosus s.l. isolates in Uzbekistan, and the results showed that G1 and G3 (E. granulosus s.s.) were the predominant genotypes among isolates from humans, sheep, and the definitive hosts (dog and jackal), and only 1 isolate was determined to be the G4 genotype (E. equinus).
The E. granulosus s.s. G1 and G3 genotypes are much more closely related to each other than to any other known genotype. Therefore, the sequences of cox1 and nad1, which are most commonly used for typing, were too short to clearly identify our isolates as G1 and G3 [24,25]. Recently, successful differentiation of the G1 and G3 genotypes was achieved by using near-complete mitochondria genome sequences (11,443 bp) [10,25]. However, we successfully classified our isolates as G1 and G3 by using only the cox1 sequences (and not nad1) along with confirmed G1 and G3 genotype reference sequences. We used the cox1 gene region from the near complete mitochondria genomes, whose genotypes were confirmed in a previous study [25]. The complete mitochondrial genome sequences have more phylogenetic information than the short sequences used in this study. However, if confirmed G1 and G3 genotype sequences are used as reference sequences for E. granulosus s.s. genotyping, we can obtain results similar to those utilizing complete mitochondria genome sequences.
We confirmed the genotypes of the isolates from jackal, sheep, and dog as G1 and G3 and 1 isolate from human as G4 (Fig. 1, Supplementary Fig. S1). In this study, we obtained 3 notable results. First, we found that the transmissibility of the parasite in intermediated hosts might be determined by its genetic background. In this study, the 10 isolates from lungs were grouped with genotype G1. E. granulosus s.l. is known to form hydatid cysts in certain organs, especially in the liver and lungs [26]. Previous studies also showed an interesting result; 68% of the E. granulosus cysts in infected pigs were found in the lungs, whereas 73% of the cysts in cattle were found in the liver [27]. This markedly different organ distribution between the 2 species might be consistent with different genotypes circulating in different species [11]. Although we have no conclusive evidence to support the hypothesis that some genotypes are better able to transfer to the lung, in the present study, all isolates from the lung were grouped with the G1 genotype. To test this hypothesis, additional analyses using many more confirmed G1 and G3 sequences are needed. Second, infections by Echinococcus eggs from sheep and humans in the definitive hosts (dog and jackal) might be uncommon recently because the genetic distance between the isolates from jackal and dog and the isolates from sheep and human were great, although 2 isolates from human were very closely related to the jackal isolate. This result may determine which genotypes are the dominant cause of EC in Uzbekistan. The final notable finding is the first report of a human E. equinus infection. The biological differences in E. equinus when compared to E. granulosus s.l. have been confirmed [28]. This genotype was first detected as a parasite in a British horse, and it differed markedly in morphology compared to E. granulosus of sheep origin [29]. It seems to be a highly specific parasite of Equidae (horses, donkeys, and zebras), as all cases involving the G4 genotype, except one, were in the Equidae. Boufana et al. [30] first reported E. equinus infection in a primate (lemur), and in Jordan, although most dogs were infected with G1, a single sample was shown to be the most similar to E. equinus (G4) using RAPD-PCR [31]. These results suggest that E. equinus might have some potential for human infection in Middle Asia. We are currently conducting epidemiological investigations in Uzbekistan to determine if G4 genotype infections occur in horses or dogs in the region.
In conclusion, we confirmed that E. granulosus s.l.-infected jackal, sheep, and dogs may be associated with human infections. The results of this study will contribute to epidemiological surveys of E. granulosus s.l. in Uzbekistan, which will be helpful for improving public health and the economy.