Ancient DNA (aDNA) analysis on archaeological samples is very important to know the change in genetic traits of parasite species through history. The earliest aDNA studies on Ascaris spp. were those of Loreille et al. [1] and Leles et al. [2]. In the reports, Ascaris aDNA could be successfully obtained by amplification with the archaeological samples, which later became the fundamental technique for phylogenetic analysis of the species. However, though the reports were a valuable enough for concerned researches on Ascaris lineage, we also admit that similar studies should be done more on the other archaeological samples because the quantity of Ascaris aDNA data is still insufficient for understanding its complete evolutionary history. In this regard, the paleoparasitological studies on archaeological samples from Joseon tombs in Korea are very important, by which the existing pool of Ascaris aDNA in East Asia could be expanded.
Actually, we previously reported a number of paleoparasitological studies with the samples from archaeological sites in South Korea [3-11]. Of them, the amplification of Ascaris aDNA could be available from a Joseon mummy case [12]. As its preservation status was quite good even after several hundred years of burial, aDNA could be extracted from ancient coprolites successfully, and then be amplified by PCR. Fortunately enough, recently, from another Joseon tomb found in Seoul City, once again we discovered the coprolite in which many ancient Ascaris eggs were still remained. The molecular analysis on the Ascaris aDNA in the sample, if it will be done successfully, can be a valuable addition to the existing genetic pool of the ancient Ascaris. We therefore tried to extract, amplify, and analyze Ascaris aDNA from newly collected ancient sample. It should be very suggestive to a clearer understanding of the Ascaris infection prevalent among pre-modern East Asian people.
In 2007, archaeologists in Hangang Institute of Cultural Heritage (Seoul, South Korea) found the Joseon tombs in Sinnae-Dong of Seoul Metropolitan City [5]. They discovered 2 coffins in the same burial pit, which should be those for a wife and a husband. Of them, the husband's coffin (SN2-19-1) was examined by us. Most of dead person's soft tissues were already mummified when we opened the coffin. We collected soil sediments from 8 different areas (Samples A to H) on the basal plate of coffin. From the surface of the sacrum, we also collected precipitates upon it (Sample sacrum). The outcome of microscopic examination on this case was already reported by us [5]. Briefly, A. lumbricoides eggs (64.0±2.3 μm ×52.5±1.7 μm) were discovered in the samples of E, F, H, and sacrum. The estimated average number of Ascaris eggs was 6,408.3 per gram.
Using the Ascaris egg positive samples, our aDNA analysis was performed. Above all, to make our aDNA work much authentic, we followed the criteria of authentication [13,14]. During all the lab procedures, we wore protection gloves, masks, gowns, and head caps. We did all experiments in our aDNA lab facility that is set up in accordance with the suggestions of Hofreiter et al. [13] or Willerslev and Cooper [14]. We could have ruled out the possibility of contamination by animal (e.g., pigs) or human feces from the current case, confirming the authenticity of origin of parasite eggs we identified in the samples.
Sediment samples including parasite eggs were treated with 1 ml lysis buffer (EDTA 50 mM, pH 8.0; 1 mg/ml of proteinase K; SDS 1%; 0.1 M DTT) at 56˚C for 24 hr. DNA extraction was performed by phenol/chloroform/isoamyl alcohol (25:24:1) method. Primers for Ascaris cytochrome b (cyt b) or Ascaris 18S ribosomal RNA (rRNA) gene were prepared, following the methods of Loreille et al. [1] and Oh et al. [12]. The primers used for amplification of mitochondrial cyt b fragment were Asc1 (5ʹ-GTT AGG TTA CCG TCT AGT AAG G-3ʹ) and Asc2 (5ʹ-CAC TCA AAA AGG CCA AAG CAC C-3ʹ). 18S rRNA region was amplified by Asc 6 (5ʹ-CGA ACG GCT CAT TAC AAC AG-3ʹ) and Asc 7 (5ʹ-TCT AAT AGA TGC GCT CGT C-3ʹ); Asc 8 (5ʹ-ATA CAT GCA CCA AAG CTC CG-3ʹ) and Asc 9 (5ʹ-GCT ATA GTT ATT CAG AGT CAC C-3'); Asc 10 (5ʹ-CCA TGC ATG TCT AAG TTC AA-3ʹ) and Asc 11 (5ʹ-CAR AAA WTC GGA GCT TTG GT-3ʹ). The amplicon sizes were 142 (Asc1/ Asc2), 123 (Asc6/Asc7), 99 (Asc8/Asc9), and 147 (Asc10/Asc11) base pairs (bps), respectively.
DNA amplification was done with 20 μl reaction mixture containing 1× High Fidelity PCR buffer, 2 mM MgSO4, 200 μM dNTP mixture, 2 unit of Platinum® Taq DNA Polymerase High Fidelity (Invitrogen, Hercules, California, USA), 10 pmol of each primer, and 1 mg/ml of BSA (New England Biolabs, Beverly, Massachusetts, USA). PCR conditions were as follows: pre-denaturation at 94˚C for 10 min; 50 cycles of denaturation at 94˚C for 45 sec, annealing at 50˚C for 45 sec, extension at 72˚C for 45 sec, and final extension at 72˚C for 10 min. In our agarose gel electrophoresis, the amplified products showed specific bands for Ascaris 18S rRNA or cyt b mitochondrial genes whereas negative controls (extraction controls) did not exhibit any amplified bands (Fig. 1).
Cloning and sequencing was also done for the amplified PCR products. Briefly, after aDNA in amplified bands was extracted by QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany), bacterial transformation with amplified DNA product was done using pGEM-T Easy Vector system (Promega, Madison, Illinois, USA). Transformed bacteria were grown in agar plate containing ampicillin (50 μg/ml), 0.5 mM IPTG, and XGAL (40 μg/μl) for the next 14 hr. After selected colonies were grown once again in LB media for 12 hr, plasmid was harvested using QIAprep spin miniprep kit (Qiagen). Sequencing for each clone was done by ABI Prism 3100 automatic sequencer (Applied Biosystems, Foster City, California, USA) with ABI Prism BigDye Terminator Cycle Sequencing Ready Reaction Kit (Applied Biosystems).
We repeated cloning and sequencing several times for each amplified product, to get the consensus sequences. By these trials, 30 clone sequences were successfully obtained from 18S rRNA gene while 12 clones were from mitochondrial cyt b gene. The pairwise and multiple sequence alignments were done by Clustal W implemented in MEGA6 [15,16]. Using the aligned clone sequences, we obtained consensus sequences for each gene (data not shown).
Next, the consensus sequences were compared to those available in GenBank by NCBI/BLAST tools [17]. Web browser module and Alignment Explorer in MEGA6 were used for retrieving sequences homologous to those of interest from National Center for Biotechnology Information (NCBI) GenBank database. Ascaris mitochondrial cyt b gene sequence obtained from SN2-19-1 were 99% identical to the ancient Ascaris sequences reported in the study of Oh et al. [12] (GU339224.1). It also showed similarities to cyt b genes of A. lumbricoides (99%, KF798183.1) and A. suum (97%, HQ704901.1). However, Baylisascaris (KC797002.1) exhibited only 94% similarity to ours (Fig. 2).
To estimate the evolutionary divergence between cyt b gene sequences, the numbers of base substitutions per site from between sequences were counted for 12 nucleotide sequences. Analyses were conducted using the maximum composite likelihood model [18]. Pairwise distances between sequences obtained by MEGA6 [16] are available in Table 1. The pattern of distances between each taxon is similar to that of sequence similarities seen in Fig. 2.
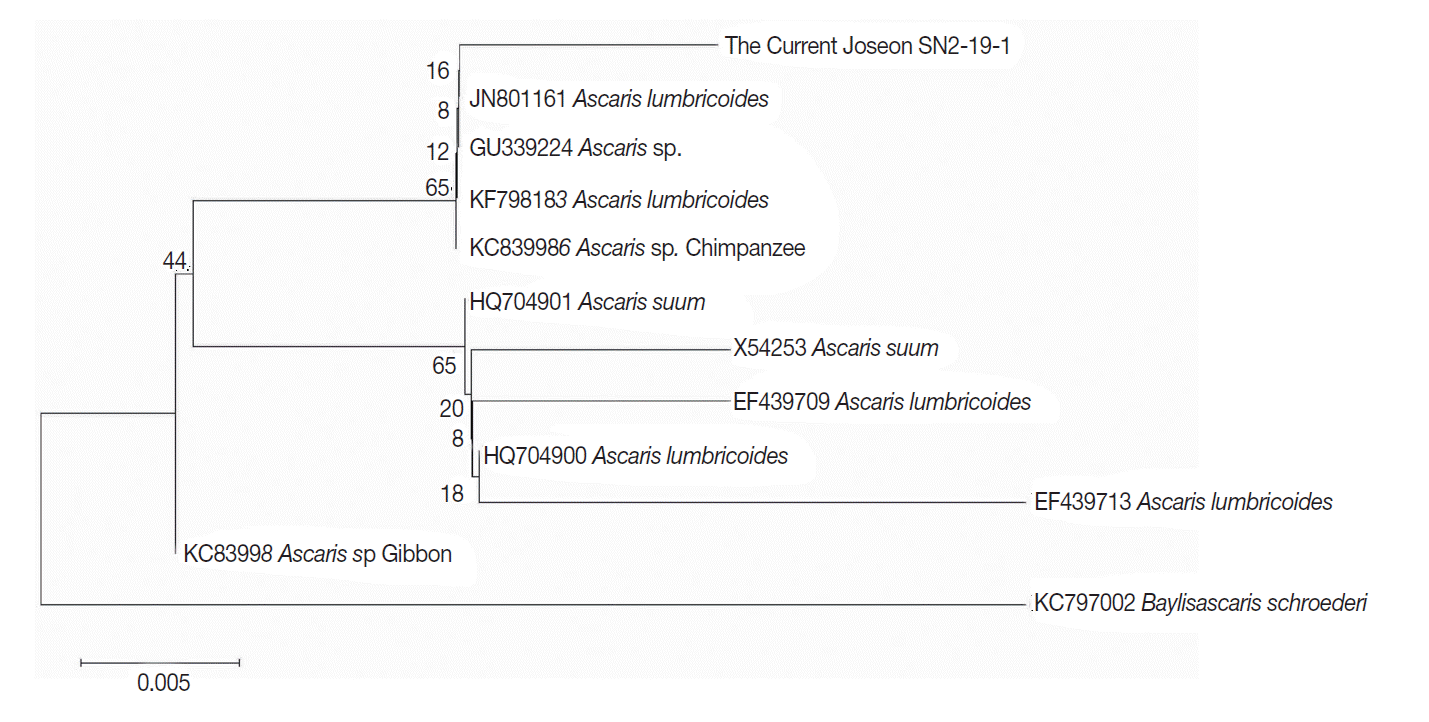
The evolutionary relationship of cyt b gene among taxa was inferred by the Neighbor-joining (NJ) method [19] in MEGA6 [16]. Selected parameters were ‘gamma distributed with invariant sites (G+I)’ for rates among sites, and partial deletion for gaps/missing data treatment. All positions with less than 95% site coverage were eliminated. Fewer than 5% alignment gaps, missing data, and ambiguous bases were therefore allowed at any position. To estimate the reliability of the tree, we tested bootstrap with MEGA6 [16]. The number of bootstrap replicates was 1,000 [20]. NJ tree for cyt b gene sequences with bootstrap values is seen in Fig. 3.
In the NJ tree, we could find 2 different clusters showing relatively higher in bootstrap values (65% for both) (Fig. 3). Cyt b sequence of the current Joseon SN2-19-1 belonged to the cluster in which A. lumbricoides sequences were mainly found. In another cluster, however, we found both A. lumbricoides and A. suum sequences. Considering that bootstrap values are not so high (below 70%) for each node of branches (Fig. 3), we need more studies on ancient and contemporary Ascaris to accept the pattern of current dendrogram seriously.
Meanwhile, 18S rRNA gene sequence obtained in this study was not only specific for Ascaris. It was 100% identical to ancient Ascaris DNA of Oh et al. [12] (GU339223.1), and to the other A. suum and A. lumbricoides sequences available in GenBank. We also noted that the 18S rRNA sequence of genus Baylisascaris exhibited 100% match with that of Ascaris (Fig. 4). In general, 18S rRNA genes have been used for the species identification, especially for studying evolutionary relationship between different species [1,12,21]. As far as Ascaris is concerned, however, we agree with Søe et al. [22]'s opinion that 18S rRNA gene is not good for molecular differentiation of Ascaris from other nematodes. Considering that mitochondrial cyt b in this study showed much significant information on molecular differentiation of each nematode species than 18S rRNA gene did, our future studies should concentrate more upon the Ascaris mitogenome, especially on its hypervariable regions.
Most Ascaris sequences in GenBank currently are uploaded as separate species such as A. lumbricoides and A. suum. Besides low divergence in morphology [23-26], A. lumbricoides and A. suum could not be differentiated easily by any of molecular markers either [21]. Researchers therefore claimed nowadays that both are not different species at all, but are a single species only infecting the different hosts by chance: humans and pigs [27-29]. Actually, considering that Ascaris isolated from chimpanzee (KC839986.1) and gibbon (KC839987.1) did show very similar cyt b sequence to those of A. lumbricoides (Fig. 2), Ascaris as a single parasite species might infect much broader range of host animal species than expected.
Taken together, the parasite Ascaris spp. are of great public health concern because it still infects 1.2 billion people worldwide [30]. To obtain the solid evidence for hypothesis about Ascaris evolution, however, much more data should be added to the extant genetic pool of Ascaris [27]. This is also true for Ascaris aDNA because the sequences from several-hundred to -thousand year old samples can provide valuable information on molecular evolution that could not be easily obtained from the contemporary Ascaris samples. More studies on Ascaris aDNA can enrich our knowledge on genetic history of the Ascaris that were prevalent among pre-modern Korean and East Asian peoples.