INTRODUCTION
Malaria is a worldwide public health problem because it is a medical emergency with a high risk of mortality [1]. Plasmodium falciparum causes the most dangerous form of malaria to humans [2]. It also exhibits resistance to antimalarial drugs [3]. The resurgence of malaria is partly attributed to wide distribution of drug-resistant strains [4]. The enzyme protein farnesyltransferase is an ideal drug target for P. falciparum [5]. Farnesyltransferase is an enzyme that belongs to the prenyltransferase group [6]. It catalyzes the posttranslational modification of intracellular signal transduction proteins by transferring the farnesyl residue of farnesyl pyrophosphate [7]. The cysteine residue, which is the transferring farnesyl residue, belongs to the carboxy-terminal consensus sequence called the CAAX motif [7]. The targets of farnesyltransferase include members of the Ras superfamily, which are critical to cell cycle progression [8]. For this reason, inhibitors of farnesyltransferase have effects on anti-cancer drugs and anti-parasitic agents [7]. Therefore, FTIs has been recommended as a novel antimalarial drug [9].
Mitochondria have been shown to play a key role in the apoptotic process through several signaling pathways. Mitochondria control the apoptosis at several levels such as maintenance of ATP production and mitochondrial membrane potential (ΔΨm) and permeability for the release of certain apoptogenic factors from the intermembrane space into the cytosol [10]. Malaria parasites have very small mitochondrial genomes, including 3 encoded proteins, and highly fragmented ribosomal RNAs [11]. The mitochondria of P. falciparum enable it to adapt to the host morphologically and physiologically [12]. The energy metabolism of P. falciparum differs from that of the other mammalian hosts [12]. P. falciparum has a simplified metabolism with biosynthetic pathways [11]. The mitochondria of P. falciparum do not exhibit full oxidation of glucose to fuel mitochondrial ATP synthesis in the malaria parasite [11]. The mitochondrial electron transport chain (mETC) is important for pyrimidine biosynthesis [11]. The mitochondria of P. falciparum are promising targets for antimalarial drugs because the energy-transducing pathways are distinct in the malaria parasite [12]. Thus, the inhibition of mitochondria has been suggested as a novel strategy for malaria extermination. However, the relationship between FTIs and mitochondria function of P. falciparum has not yet been investigated.
In this study, the effects of FTI (Zarnestra, R115777) on the mitochondrial function of P. falciparum were investigated under in vitro condition. In addition, to compare the drug target, after treating chloroquine and atovaquone in vitro, the copy number of mtDNA-encoded cytochrome c oxidase III was measured for each case. The function of mitochondria was examined by staining with the mitochondrial membrane potential (ΔΨm)-sensitive dyes and MitoTracker.
MATERIALS AND METHODS
Chemicals
FTI was purchased from Selleck (Zarnestra, R115777, Houston, Texas, USA). Chloroquine and atovaquone were purchased from Sigma-Aldrich (St. Louis, Missouri, USA).
Parasite culture
P. falciparum (3D7 strain) was maintained in human erythrocytes (blood group O+) in RPMI 1640 (buffered with 25 mM HEPES and 25 mM NaHCO3) supplemented with 10 μg/ml gentamycin and 370 μM hypoxanthine under a gas mixture containing 5% CO2, 5% O2, and 90% N2 at 37˚C in an incubator. Parasites were diluted with 9.5 ml of culture medium containing 0.5 ml of sufficient non-infected human erythrocytes to yield a final hematocrit of 5% and a parasitemia of 0.5% [13].
Giemsa staining
A thin blood smear was fixed with 99% methanol for 5 min. After fixing the slide, it was dried for 1 min to 2 min. A diluted Giemsa solution (Merck, Darmstadt, Germany) was prepared prior to staining. The slide was immersed in freshly prepared Giemsa stain solution for 25 min, rinsed with tap water, and dried. The stained malaria parasites were examined using a microscope with a 100× oil immersion objective lens.
Synchronization of P. falciparum
Synchronized ring-form cultures were tested to investigate the antimalarial effects. Sorbitol (5%) was dissolved in distilled water, and passed through a Millipore filter of 0.22 μm pores. Sorbitol (9 ml) was added to 1 ml of the culture pellet, and incubated for 5 min in room temperature. The supernatant was removed by centrifugation, and the pellet was washed 3 times in complete media. P. falciparum was determined to be free of schizonts by microscopic examination of the Giemsastained specimen [14]. After synchronization, ring form of P. falciparum were treated with each drugs for 48 hr. The infection rate in synchronized P. falciparum was approximately 1.8-2.0%.
MitoTracker labeling
To measure the mitochondria mass, MitoTracker Green was treated in malaria-infected RBCs after FTI R115777 treatment for 48 hr. The 9 ml of supernatant of malaria-infected RBCs was removed by centrifuge and malaria-infected RBCs were incubated with 1 mM MitoTracker stock solution for 30 min at 37˚C. After staining with a MitoTracker dye, the malaria-infected RBCs were washed in incomplete media. Fluorescence imaging techniques were employed with the excitation and emission wavelength bands centered at 490 and 516 nm, respectively. Fluorescence was measured by a CytoFluor fluorescence multiwell plate reader (Tecan USA, Durham, North Carolina, USA). Fluorescence signal intensities were evaluated using Image J software (ImageJ 1.46r software, U.S. National Institutes of Health, Bethesda, Maryland, USA).
Determination of mitochondrial membrane potential (ΔΨm)
Variations in ΔΨm were analyzed using 5, 5´, 6 ,6´-tetrachloro-l, l´, 3, 3´-tetraethylbenzimidazolcarbocyanine iodide (JC-1) (Molecular Probes, Invitrogen, Eugene, Oregon, USA), which was stained as previously described [15]. Malaria parasite infected RBCs treated with FTI R115777 for 48 hr were incubated with JC-1 reagent for 20 min at 37˚C. The supernatant was removed by centrifugation, and the pellet was washed 3 times in Cell-Based Assay buffer (Molecular Probes, Invitrogen). Malaria-infected RBCs were analyzed using a CytoFluor fluorescence multiwell plate reader. For healthy conditions, JC-1 forms J-aggregates in excitation and emission wavelengths of 560 and 595 nm, respectively. For apoptotic or unhealthy conditions, JC-1 exists as a monomer at 485 and 535 nm. Fluorescence signal intensities were measured using Image J software.
Determination of oxygen gradients
P. falciparum was treated with FTI R115777 for 48 hr after sorbitol synchronization. The sensor foil (SF-RPSU4, PreSens Precision Sensing GmbH, Regensburg, Germany) was calibrated by placing a drop of sodium dithionate (Na2S4O4), and recording the image [16]. The oxygen concentration was evaluated using VisiSens Analytical 1 software. After recording the calibration value, the sensor foil was sterilized by 70% ethanol, and washed with autoclaved distilled water several times. Malaria-infected RBCs were placed on the oxygen-sensitive sensor foil, and data were read using a detector unit.
Detection of reactive oxygen species (ROS) content by dichlorofluorescein (DCF)
To measure ROS activity, an OxiSelect ROS assay kit (Cell Biolabs, San Diego, California, USA) was used as previously described [17]. The assay uses the diffusion of 2´,7´-dichlorodihydrofluorescin diacetate (DCFH-DA) into cells. DCFH-DA is deacetylated by cellular esterases to non-DCFH and rapidly oxidized to DCF by ROS. In brief, malaria-infected RBCs were pre-incubated with DCFH-DA (100 μM) for 20 min before adding FTI R115777. A CytoFluor Fluorescence Multiwell Plate Reader (Tecan USA) was used to determine ROS production by comparing the results with a predetermined DCF standard curve (excitation=480 nm, emission=520 nm). Fluorescence signal intensities were measured using Image J software.
Isolation of mitochondrial DNA
P. falciparum was maintained for 48 hr in drugs for treatment after sorbitol synchronization. The mtDNA was isolated using mtDNA Extractor CT kits (Wako pure Chemical Industries, Ltd., Osaka, Japan) following the manufacturer’s instructions [18]. Malaria parasites were homogenized in ice-cold homogenization buffer provided in the kit. The homogenate was centrifuged at 1,000 g and 4˚C for 1 min. The supernatant was collected and centrifuged at 10,000 g and 4˚C for 10 min. The mtDNA extraction solutions in the kit were added to the pellet. mtDNA was precipitated by the addition of 300 μl of sodium iodide solution and 500 μl of isopropanol. The mtDNA concentrations were measured using Nano-Drop (Thermo Scientific, Wilmington, Delaware, USA).
mtDNA amplification
All oligonucleotide primers were used as described in Cunha et al. [19]. The sequences of cytochrome c oxidase III (GenBank accession nos. GI8346992 and M76611) expressed from P. falciparum used in this study were Pf1 (5´-CCTGCATTAACATCATTATATGGTACATCT-3´) and Pf2 (5´-GATTAACATTCTTGATGAAGTAATGATAATACCTT-3´) [19]. PCR was conducted using Accupower PCR premix (Bioneer, Daejeon, Korea). The equivalent amount of the mtDNA were used. All procedures were performed according to the manufacturer’s instructions. The amplification conditions were as follows: 1 denaturation cycle at 96˚C for 10 min; 30 cycles at 95˚C for 1 min and 60˚C for 5 min; and a final extension at 60˚C for 1 hr. All amplifications were performed in a MyGenie 96 Gradient Thermal Block (Bioneer). The PCR products were resolved with standard agarose gel electrophoresis, and visualized using ethidium bromide staining under UV light.
RESULTS
Growth inhibitory activity of FTI R115777
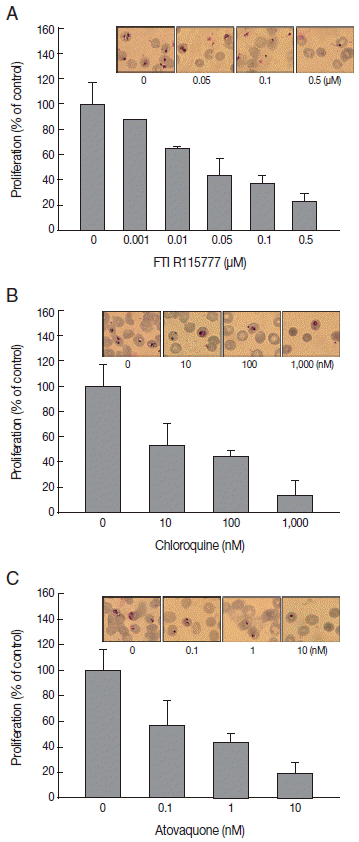
To elucidate the antimalarial effects of FTI R115777, P. falciparum was treated with the drug for 48 hr. Before treating drugs, parasitemia in synchronized P. falciparum was approximately 1.8-2.0%. Parasitemia after drug treatment was 13.4±0.3% in the control group. FTI R115777 inhibited the proliferation of malaria parasites in a dose-dependent manner. The IC50 for FTI R115777 for P. falciparum 3D7 was 11.48 nM in vitro. FTI R115777 (100 nM) inhibited the growth of parasites by 65.2±1.8% (Fig. 1A). Malaria parasitemias became 5.8%, 5,6%, and 3.0% after addition of 0.05, 0.1, and 0.5 μM of FTI R115777, respectively. The high-dose FTI R115777-treated P. falciparum group exhibited distinctive morphological features, such as pyknosis form, compared with the control group (Fig. 1A). In addition, we treated atovaquone, known to target the mitochondria, and chloroquine that does not have a specific mitochondrial effect [20,21]. Chloroquine and atovaquone were observed to inhibit P. falciparum infection as coincided with Fig. 1B and 1C. Malaria parasitemias became 6.8%, 5.7%, and 1.7% after addition of 10, 100, and 100 nM of chloroquine, respectively (Fig. 1B). When 0.1, 1, and 10 nM of atovaquone was treated, parasitemia was 7.7%, 5.9%, and 1.2%, respectively (Fig. 1C). These results showed the antimalarial effect of FTI R115777, cloroquine, and atovaquone.
Maintenance of mitochondrial membrane potential ΔΨm
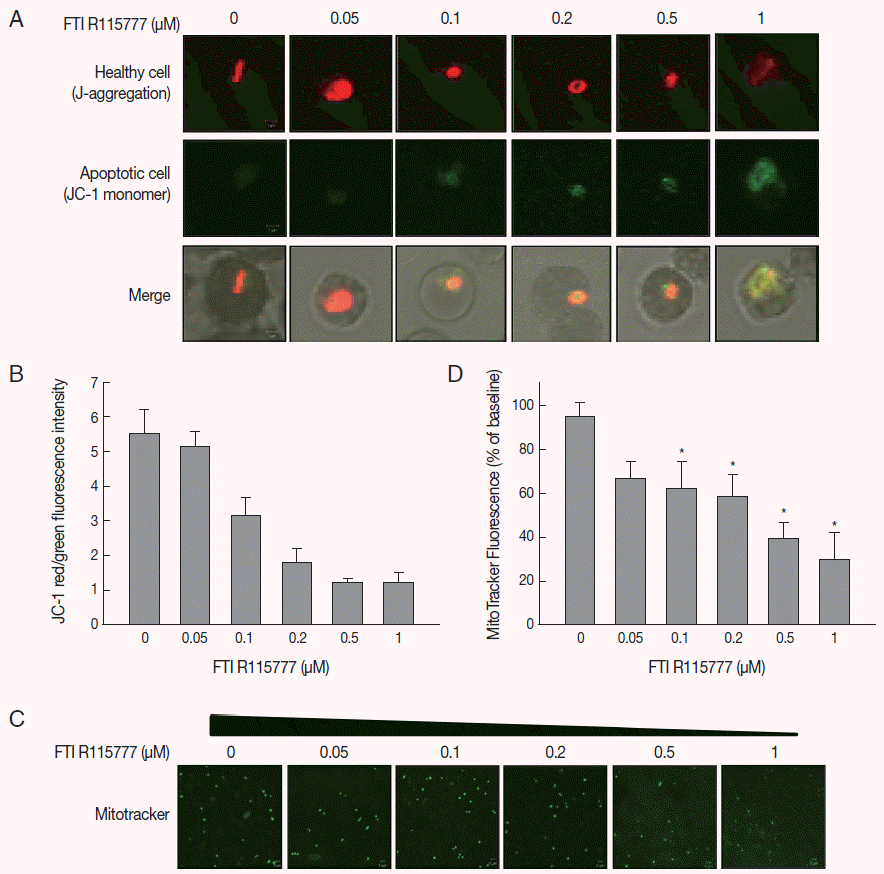
Given that FTI R115777 suppresses the proliferation of P. falciparum, we investigated whether this action works specifically through the mitochondria. As shown in Fig. 2A and B, the low-dose FTI R115777-treated group exhibited strong red fluorescence, which indicated a healthy condition. By contrast, the high-dose R115777-treated group exhibited green fluorescence because of low ΔΨm compared with that of the control group. These results showed that mitochondrial depolarization was induced by FTI R115777.
Intensity of MitoTracker green fluorescence in FTI R115777-treated malaria parasites
To investigate the effect of FTI R115777 on the mitochondrial function of P. falciparum, MitoTracker green fluorescence was stained in vitro. MitoTracker labels the mitochondria in live cells using ΔΨm, and can be used for multiple labeling and measuring of the depolarized mitochondrial mass [22,23]. The cells stained by MitoTracker were observed under fluorescence microscopy, and the fluorescence signal intensities were measured using Image J software. Fig. 2C and D clearly show that the MitoTracker green fluorescence signal decreased as the dose of FTI R115777 increased. This result indicated that FTI R115777 can be used as an effective target to mitochondria.
Oxygen concentration of FTI R115777-treated malaria parasites
To examine the mitochondrial activity, extracellular oxygen concentrations were measured via the ratiometric read-out of the sensor foil by VisiSens devise. The oxygen gradient of FTI R115777-treated P. falciparum were shown on the sensor foil (Fig. 3A). As shown in Fig. 3B, the oxygen concentration of malaria-infected RBCs was higher than that of the non-infected RBCs (P<0.05). However, the average oxygen concentration started to decrease slightly from the dose of 0.1 μM to 1 μM FTI R115777. This result implied that depolarization was mainly mediated by other contributors rather than oxygen consumption.
Effect of FTI on the intracellular ROS level
We investigated whether FTI R115777-treated P. falciparum was linked to mitochondrial ROS generation. The DCFH-DA-measurable ROS induced by FTI R115777 in the malaria parasites was measured. ROS levels notably increased in malariainfected RBCs than those in non-infected RBCs. The intracellular ROS levels increased as the dose of FTI R115777 increased (Fig. 3C). This result indicated that the FTI R115777-induced intracellular ROS could sufficiently disrupt the mitochondrial functions. The specificity of FTI R115777 could be attributed to the selective interaction with malarial mitochondria. This interaction may specifically generate ROS in malarial mitochondria, which are key factors that enable the manifestation of the antimalarial drug effect of FTI R115777.
Copy number of cytochrome c oxidase III in FTI R115777-treated P. falciparum
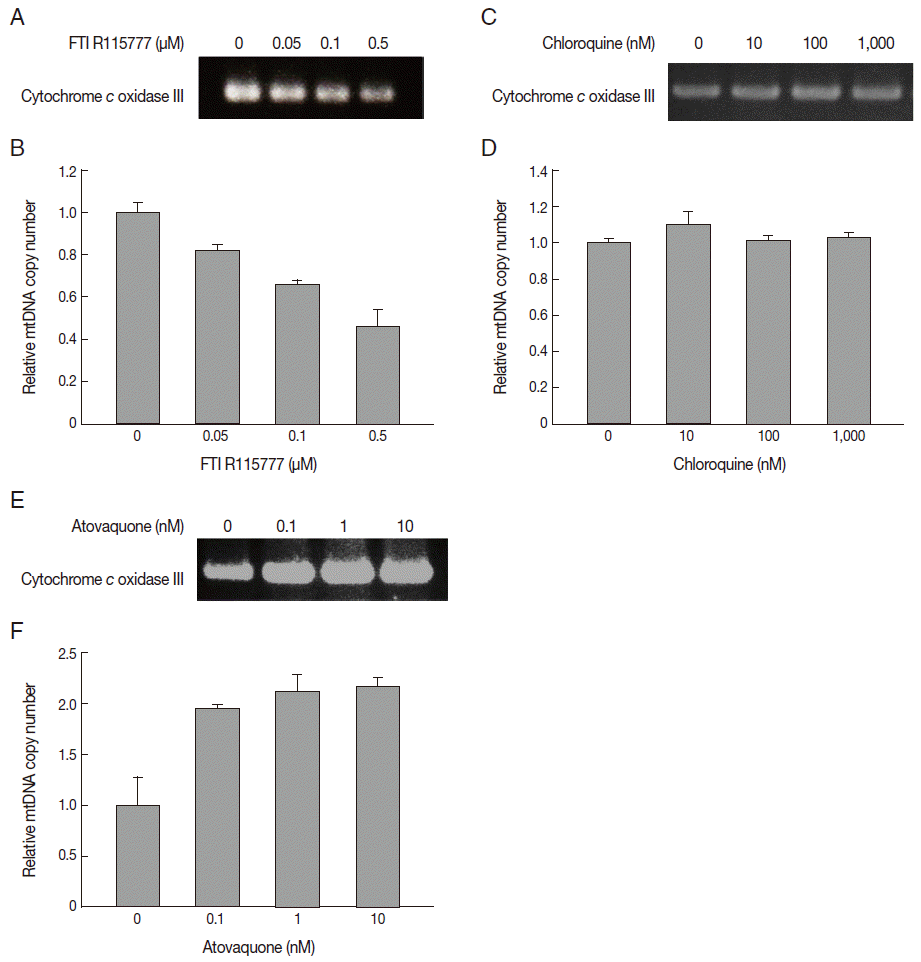
In this study, we measured the copy number of mtDNA-encoded cytochrome c oxidase III in FTI R115777-treated P. falciparum. FTI R115777 was observed to reduce the copy number of mtDNA-encoded cytochrome c oxidase III. The contents were dependent on the dose of FTI R115777, as shown in Fig. 4A and B. Atovaquone and chloroquine have been used as mitochondrial specific inhibitor and general parasite inhibitor, respectively. The copy number of mitochondrial DNA-encoded cytochrome c oxidase III was compared for chloroquine and atovaquone-treated P. falciparum. Chloroquine did not affect the copy number of mtDNA-encoded cytochrome c oxidase III, while atovaquone increased the corresponding copy number (Fig. 4C-F). These results suggested that mtDNA-encoded cytochrome c oxidase III of P. falciparum can be a potential target of FTI R115777.
DISCUSSION
FTIs are selective inhibitors of intracellular farnesyltransferase (FTase). FTIs are substrates for prenylation via FTase, including Ras, Pho-B, Rac, Pheb, nuclear lamins, and centromeric proteins, which interact with microtubules to promote the completion of mitosis [24]. FTase inhibition prevents the posttranslational processing of Ras proteins, which exert effects on multiple mechanisms of cellular survival, including angiogenesis, cellular adhesion, and mitosis [24]. FTI R115777 (Zarnestra, Tipifarnib) is a nonpeptidomimetic FTI with clinical ability in some cancers, such as metastatic breast cancer, leukemia, and metastatic glioma [25]. A previous study demonstrated the cytotoxicity of FTIs to P. falciparum. The IC50 of FTIs ranges from 0.7 nM to 1,000 nM in vitro [5]. Thus, FTI is a promising antimalarial agent. However, the target of FTIs in organelles such as mitochondria has not yet been investigated.
The mitochondrion in malaria parasites has a unique branched tricarboxylic acid metabolism without any activation of a pyruvate dehydrogenase. It does not show ATP production in blood stages of the parasite. A previous study has shown that mitochondrial electron transport chain of P. falciparum is involved in pyrimidine biosynthesis enzyme dihidroorotate dehydrogenase in blood stages [26]. In addition, it generates an electrochemical proton gradient across the inner membrane of mitochondrion that is utilized by the ATP synthase to drive the synthesis of ATP. The functional consequence of ATP synthase is interesting, because oxidative phosphorylation is not a major source of ATP in P. falciparum blood stages. Therefore, mitochondria of Plasmodium could be a target for antimalarial drugs [11]. Mitochondrial dysfunction can induce apoptotic pathways [27]. The mitochondria control apoptosis at several levels, such as maintenance of ATP production and ΔΨm and mitochondrial membrane permeability, to release certain apoptogenic factors from the intermembrane spaces into the cytosol [10]. Thus, ΔΨm is a governing parameter of mitochondrial functions and cell health [27]. Changes in ΔΨm are observed by staining with JC-1, a lipophilic cationic dye that selectively penetrates the mitochondria. This study shows that mitochondrial depolarization were induced by FTI R115777 dose dependently (Fig. 2).
The membrane potential generated by ATPase and mitochondrial electron transport chain (mETC) may be necessary because of the low oxygen consumption rate of P. falciparum. The decrease in ATP utilization inhibits oxygen consumption. Moreover, the respiration rate increases when ATP synthesis increases, thereby leading to the concept of respiratory control by ADP phosphorylation [28]. Oxygen is a terminal acceptor in the mitochondrial electron transport chain (mETC). Thus, the intracellular oxygen concentration has critical functions in physiological and pathological processes [29]. In a previous study, the extracellular and intracellular oxygen concentrations were assumed to be similar [29]. In the present study, we observed that the oxygen concentration of malaria-infected RBCs was higher than that of the non-infected RBCs. However, FTI R115777 did not affect the oxygen level in malaria-infected RBCs (Fig. 3A, B). The mitochondria depolarization by FTI R115777 could be affected by other factors than oxygen consumption.
Mitochondria are crucial cellular energy-transducing organelles that function as a large source of ROS. ROS production can be controlled by ΔΨm. Dissipation of ΔΨm leads to uncoupling of the electron transport chain [30]. ROS are required for mosquitoes to induce effective immune responses against Plasmodium [31]. Therefore, mitochondrial ROS are important activators in the immune response of the host to malaria infection [32]. The generation of ROS has been known to have a crucial function in the development of systemic complications in malaria [33]. In this study, ROS levels significantly increased in malariainfected RBCs than in non-infected RBCs. In addition, FTI R115777 induced the level of the intracellular ROS. This is true that FTI R115777-induced intracellular ROS could sufficiently disrupt the mitochondrial functions. The specificity of FTI R115777 could be attributed to the selective interaction with malarial mitochondria. This interaction may specifically generate ROS in malarial mitochondria, which are key factors that enable the manifestation of the antimalarial drug effect of FTI R115777.
The mitochondrial electron transport system can be used as a target for antimalarial drugs of P. falciparum [34]. In intra-erythrocytic malaria parasites, the mitochondrial electron transport system transfers the reducing equivalents generated by dihydroorotate dehydrogenase of the de novo pyrimidine biosynthetic pathway to ubiquinol-cytochrome c reductase [34]. The cytochrome c oxidase complex conducts the final step of electron transport in malaria parasites [35]. Plasmodium cytochrome c oxidase has the most streamlined subunit composition among eukaryotes [36]. Thus, the expression of cytochrome c oxidase represents the functions of Plasmodium. Cytochrome c is released from mitochondria early in apoptosis before mitochondrial depolarization, activation of caspases, and DNA fragmentation [37].
Atovaquone which was introduced in the early stage of antimalarial drug development, has been known to inhibit the activity of cytochrome c reductase in cholate-lysed mitochondria from P. falciparum and P. yoelii. It works on the cytochrome bc1 complex in the malarial respiratory chain [38]. The role of mitochondria of malaria parasites is to dispose electrons generated by dihydroorotate dehydrogenase which is an essential enzyme in pyrimidine biosynthesis [38]. Therefore, the inhibition of dihydroorotate dehydrogenase has been suggested as antimalarial compounds, such as atovaquone [38].
In this study, we isolated the mitochondrial DNA and measured the copy number of mitochondrial DNA-encoded cytochrome c oxidase from FTI R115777-treated P. falciparum. FTI R115777 inhibits the copy number of mitochondrial cytochrome c oxidase dose dependently. mtDNA copy number were measured after treatment of chloroqunine and atovaquone as mitochondrial non-specific or specific inhibitor, respectively. FTI R115777 and atovaquone shows differences in the copy number of mtDNA-encoded cytochrome c oxidase III. These drugs may act as a target of mitochondria through the distinguished mechanisms. To explain this different response of the copy number of mtDNA-encoded cytochrome c oxidase III by FTI R115777 and atovaquone needs to be verified through a further study in the future.
The effect of FTI R115777 on mitochondria of P. falciparum was in vitro analyzed in this study. The FTI R115777 was observed to inhibit the growth of malaria parasites effectively. It also affected the mitochondrial depolarization and ROS levels. The copy number of mtDNA-encoded cytochrome c oxidase III was significantly reduced by FTI R115777. The experimental data obtained in this study supported that FTI R115777 has strong influence on the mitochondria of P. falciparum by deteriorating the function of malarial mitochondria.