INTRODUCTION
Helminthic parasites have evolved to protect themselves from host immune systems. Infection of the host with these parasites elicits a Th2 response that is harmful to the parasite. Nevertheless, the parasites can overcome this host immune attack by various immune evasion mechanisms, including recruitment of the CD4+CD25+Foxp3+T (regulatory T, Treg) cell subset [1-3]. Upregulation of IL-4, IL-5, and IL-13 cytokine levels and the occurrence of eosinophilia are commonly seen in Trichinella spiralis-infected mammals, but these Th2 responses are downregulated by recruitment of Treg cells [3,4]. Treg cells from parasite-infected mice were found to express more Treg-cell homing receptors (CCR5 and CCR9) and activation markers (Klrg1, Capg, GARP, Gzmb, and OX40) than those from non-infected mice. Furthermore, administration of a parasiteactivated Treg cell population could repress artificially induced Th2-mediated inflammation [3].
Recently, innate immunity against helminthic parasites was shown to involve various pattern-recognition receptors like Toll-like receptors (TLRs). Thirteen mammalian TLR subtypes (10 in humans and 12 in mice) have been identified to date [5]. The receptors of the TLR family are type I transmembrane proteins with extracellular leucine-rich repeat domains and a cytoplasmic domain with significant homology to the IL-1 receptor type I called Toll/IL-1 receptor (TIR) domain. Four adapter proteins for TLR have been identified: myeloid differentiation factor 88 (MyD88), MyD88 adapter like/TIR-associated proteins (MAL/TIRAP), Toll receptor-associated activator of interferons (TRIF), and Toll receptor-associated molecule (TRAM). These adapter proteins initiate a cascade of events that initiates the production of cytokines, chemokines, adhesion molecules, and co-stimulatory molecules [6,7]. They are highly conserved proteins that specifically recognize distinct mutation-resistant pathogen-associated molecular patterns (PAMPs). MyD88 in particular is closely linked to the Th2 immunity against T. spiralis [8]. Nevertheless, it was not known which TLR exhibits elevated expression during T. spiralis muscle and intestinal stage infections or what these potential alterations could mean.
In this study, we investigated TLR expression patterns during T. spiralis infection and delineated their significance in host immune responses.
MATERIALS AND METHODS
Mice
Six-weeks old female C57BL/6 mice were purchased from Samtako Co. (Gyeonggi-do, Korea) and were housed in a specific pathogen-free facility at the Institute for Laboratory Animals of Pusan National University.
Isolation of muscle larvae
The T. spiralis strain used in this study has been maintained in our laboratory via serial passage in rats. For acquisition of larvae in the muscles, eviscerated infected rat carcasses were cut into pieces, and then digested in 1% pepsin/1% hydrochloride digestion fluid (artificial gastric juice) for 1 hr at 37˚C with stirring. Larvae were collected manually from digested muscle solution under the microscope and washed 6 times with sterile PBS containing 100 µg/ml ampicillin, 5 µg/ml kanamycin, and 50 µg/ml tetracycline. After collection, the worms were thoroughly and carefully washed 3 times with PBS in order to prevent contamination with the host material.
Muscle larva infection
Mice were orally infected with 250 infectious T. spiralis muscle larvae, and were sacrificed at 0, 1, 2, and 4 weeks after infection. After sacrifice, parasite-infected muscle tissue and the small intestine were isolated for analysis of TLR gene expression.
Preparation of excretory-secretory (ES) proteins of muscle larvae
In order to obtain ES proteins, the collected worms were introduced into 24-well plates containing serum-free RPMI 1640 medium supplemented with antibiotics (100 µg/ml ampicillin, 5 µg/ml kanamycin, and 50 µg/ml tetracycline). The culture was then maintained for 7 consecutive days at 37˚C in 5% CO2. It was confirmed that the majority of the larvae remained alive and exhibited good mobility during this time. Following centrifugation (12,000 g for 30 min), the supernatants were concentrated by pressure applied in a concentrator (Amicon, Millipore Corporation, Billerica, Massachusetts, USA) with 3 kDa pore size membranes. Various proteins (3-100 kDa and heavier) were detected using SDS-PAGE. Excessive salts were eliminated from the collected medium by HiTrap DesaltingTM (GE Healthcare, Uppsala, Sweden). Lipopolysaccharide was then depleted (endotoxin levels <0.01 μg/ml) from ES proteins using Detoxi-Gel Affinity Pak pre-packed columns (Pierce, Rockford, Illinois, USA), in accordance with the manufacturer`s instructions.
Cell culture and in vitro stimulation
In order to compare the expression levels of various chemokine and cytokine genes, mouse embryonic fibroblast (MEF) cells from TRIF knock out (KO) and MyD88/TIRAP KO C57BL/6 mouse fetuses 10 days after fertilization were used in this study. In addition, 5×105 MEF cells were plated in 24-well plates and incubated overnight at 37˚C and 5% CO2. Cells were treated with ES proteins (final conc., 1 µg/ml) for 2 hr.
RNA extraction and cDNA synthesis
Homogenized muscle tissues, small intestine tissues, or MEF cells were mixed with TRIzol reagent (Invitrogen, Karlsruhe, Germany), and RNA was extracted in accordance with the manufacturer`s protocols. The first-strand cDNA was synthesized using MMLV reverse transcriptase with oligo (dT) 18 primers (Invitrogen).
Touchdown PCR and real-time PCR
Touchdown PCR was performed using cDNA to analyze the expression levels of TLRs (TLR1-9), IL-25, eotaxin, TSLP, TGF-β, IL-10, and IFN-γ. The PCR conditions were as follows: (1) 94˚C for 5 min; (2) 19 cycles of 94˚C for 30 sec and 68˚C for 30 sec, with the annealing temperature decreasing by 0.5˚C every second cycle from 68˚C to 55˚C; (3) 19 cycles of 92˚C for 30 sec and 60˚C for 45 sec; and (4) 4˚C for 10 min. PCR products (5 μl) were analyzed with 0.7% agarose gel electrophoresis. Based on the observed gene expression levels, genes with markedly increased expression were further analyzed via real-time PCR using the iCyclerTM real-time PCR system (Bio-Rad Laboratories, Hercules, California, USA). GAPDH was used as the reference gene. The primer sequences are shown in Table 1. Amplification of all genes was performed under the following conditions: 1 min 30 sec hot start at 95˚C, followed by for 40 cycles each of denaturation at 95˚C for 25 sec, primer annealing at 50-55˚C for 20 sec, and elongation at 72˚C and 30 sec. The fluorescence of the DNA-binding dye SYBR Green was monitored after each cycle at 50-55˚C. An iCycler iQTM multicolor real-time PCR detection system (Bio-Rad Laboratories) was used for estimation of expression levels, and the Gene-x program (Bio-Rad Laboratories) was employed to determine the relative gene expression of each gene calculated as the ratio to a housekeeping gene.
RESULTS
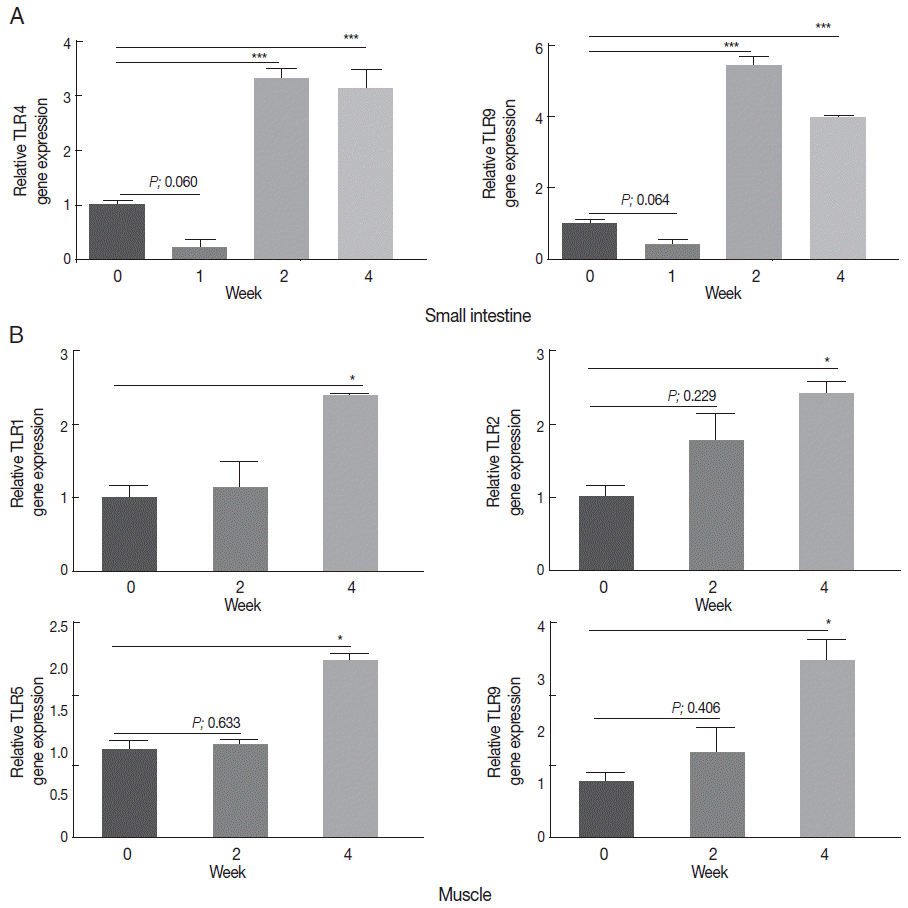
Several TLRs exhibit enhanced expression during T. spiralis infection
In order to elucidate which of the TLRs is linked to T. spiralis infection, the expression levels of 9 mouse TLR genes (TLR1-9) in the small intestine (intestinal phase) and of muscle tissue (muscle phase) were analyzed 0, 1, 2, and 4 weeks after T. spiralis infection. The results demonstrated a significant increase in TLR4 and TLR9 gene expression in the small intestine 2 weeks after T. spiralis infection, which was maintained up to 4 weeks after the infection (Fig. 1A). Although their expressions were decreased at 1 week, these are not significant. TLR9 gene expression in particular highly increased up to 6 times compared to the levels before infection. In contrast, the expression of the other TLRs was not significantly changed by the infection in the small intestine (data not shown). In the muscle, none of the TLR genes showed enhanced expression 1 week after infection, but TLR1, TLR2, TLR5, and TLR9 gene expression significantly increased 4 weeks post-infection (Fig. 1B). However, the expression of the other TLRs was not significantly changed in the muscle (data not shown).
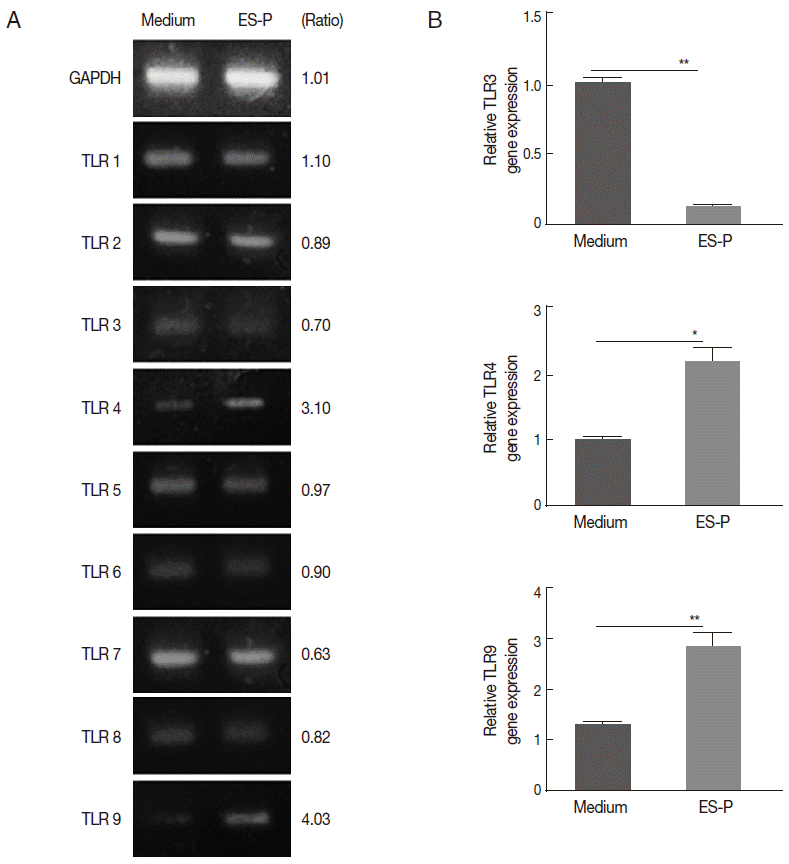
TLR4 and TLR9 gene expression was elevated by T. spiralis ES proteins in vitro
In order to determine whether ES proteins can elicit TLR gene expression, we subsequently measured the TLR expression levels in MEF cells after treatment with ES proteins. Among the TLRs, only TLR4 and TLR9 gene expression was elevated (about 3- to 4-fold) by the treatment, and TLR3 and TLR7 gene expression was slightly decreased after ES proteins treatments (Fig. 2A). In real-time RT PCR, TLR4, and TLR9 gene expression was significantly increased (about 2-3 folds) and TLR3 gene expression significantly decreased after ES protein treatment, other gene expression changes was not significant (Fig. 2B).
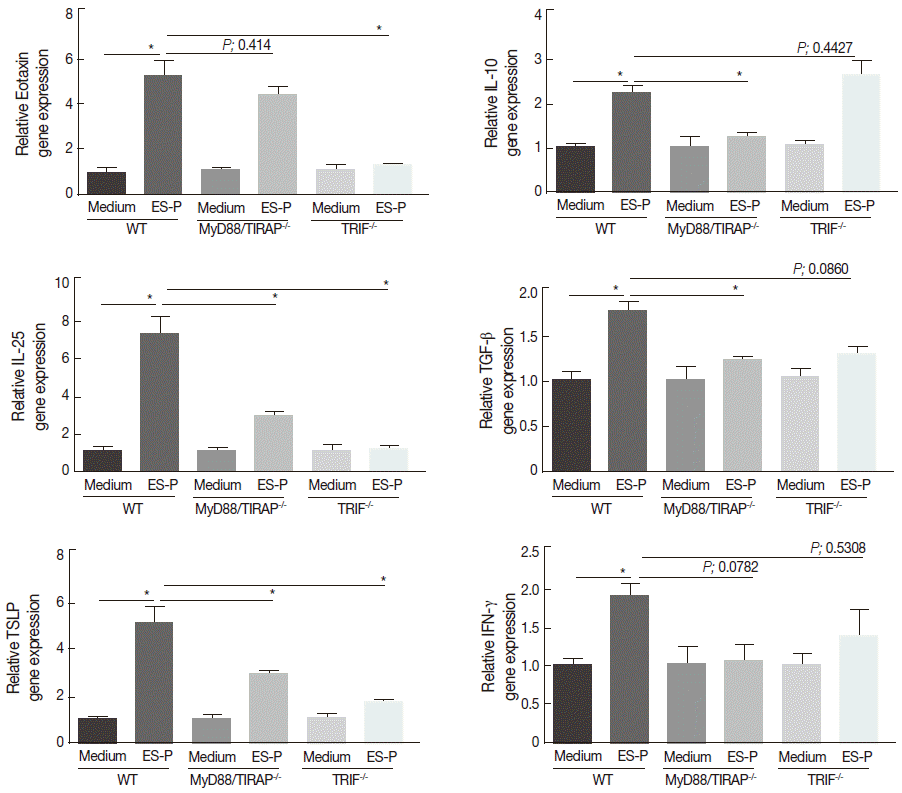
Gene expression of Treg cell-related cytokines induced by ES proteins is more closely associated with TLR9 than with TLR4
T. spiralis infection in vivo (and also ES protein treatment in vitro) can elicit Th2- and Treg cell-related immune responses. In order to determine which TLR (TLR4 or TLR9) is more important in immune responses against the parasite infection, we compared the gene expression levels of Th2 initiation chemokines (eotaxin, TSLP, and IL-25), Treg cell-related cytokines (IL-10 and TGF-β), and a Th1 cytokine (IFN-γ) in MyD88/TIRAP-/- and TRIF-/- MEF cells with those of WT MEF cells after treatment with ES proteins. The expression of Th2-related chemokine and cytokine genes decreased in both KO MEF cells, but the extent of the reduction differed (Fig. 3). IL-25 and TSLP gene expression levels were highly enhanced (about 5-8 fold) by ES proteins in WT MEF cells, but the expression levels were only slightly increased (about 3-fold) in MyD88/TIRAP-/- MEF cell, but it was not changed in TRIF-/- MEF cells. Interestingly, the eotaxin gene expression level was not changed by ES proteins treatment in MyD88/TIRAP-/- MEF cells, while it reached almost basal levels in TRIF MEF-/- cells. Nevertheless, Treg cell-related cytokine expression levels in both KO MEF cells were contrary to the Th2 chemokine and cytokine levels (Fig. 3). IL-10 and TGF-β expression levels were increased by about 2-fold in WT MEF after ES protein treatments, while they were significantly low-expressed in MyD88/TIRAP-/- MEF cells and slightly but not significantly increased in TRIF-/- MEF cells. Gene expression of the Th1-related cytokine IFN-γ was slightly but not significantly increased in both KO MEF cells (Fig. 3).
DISCUSSION
TLRs are known to provide critical defense mechanisms against bacterial and viral infections. They might also play an important role in the innate and adaptive immune responses against helminth infections, because ES proteins, glycoproteins, and microRNAs from helminthic parasites have been demonstrated to activate immune cells or an immune response related to TLRs [9-13]. T. spiralis in particular releases abundant glycoproteins throughout its life, which can exist in the phosphorylcholine form after glycan modification in the body of the parasite [14-16]. The phosphorylcholine of the parasitic helminth has been identified and was shown to bind to TLR4 [11]. Indeed, some reports have demonstrated a TLR-mediated immune suppression. Mediated by several TLRs, immune responses (activation of dendritic cells) decrease the susceptibility of effector T cells to suppression mediated by Treg [17]. Nonetheless, there is no report on the variation of TLR gene expression during the T. spiralis infection phase.
In this study, we observed that the gene expression of some TLRs was significantly increased in the intestinal and muscle phases. This finding suggests that TLRs might be closely linked to T. spiralis infection (Fig. 1), and their expression pattern was similar to that of certain cytokines after T. spiralis infection. In our previous study, Th2 cytokines and chemokines increased in mesenteric lymph node (MLN) at 1-2 weeks after T. spiralis infection, the levels of IL-10 and TGF-β in MLN and peripheral lymph nodes (PLN) were increased at 2-4 weeks after infection [4]. In order to clarify the role of TLRs in the immune response against T. spiralis, we analyzed TLR gene expression in 3 types of MEF cells isolated 2 TLR adapter protein-deficient mice and WT mice. We found that only TLR4 and TLR9 gene expression in MEF cells increased after treatment of the cells with T. spiralis ES proteins treatment (Fig. 2). TLR4 and TLR9 differ in their adapter proteins; TLR4 uses MyD88/TIRAP and TRIF, while TLR9 uses only MyD88 as [7]. Interestingly, we identified differences in Th2- and Treg-related cytokine gene expression between both adapter protein- deficient MEF cells (Fig. 3). The expression levels of these genes (IL-25, TSLP, and eotaxin) resembled basal levels in ES protein-treated TRIF-/- MEF cells. In MyD88/TIRAP MEF-/- cells, IL-25 and TSLP expression levels were significantly reduced but not to basal levels, and eotaxin expression level was not changed in. In agreement with these observations, Cho et al. [12] reported that TRIF was critical in simplex ES proteins-induced IL-6 and eotaxin (CXCL1) gene expression in Anisakis. These results show that the TRIF adapter protein is most important in Th2-related cytokine gene expression. Only TLR3 and TLR4 use TRIF as an adapter protein, but TLR3 gene expression was significantly decreased in WT MEF cells treated with T. spiralis ES proteins in this study (Fig. 2A, B). Therefore, TLR4 might be more closely associated with the Th2 response in T. spiralis infection than other TLRs.
On the other hand, IL-10 and TGF-β gene expression levels were not significantly reduced in TRIF deficiency; however, the expression of these genes was significantly reduced to basal levels in MyD88/TIRAP adapter protein deficient mice (Fig. 3). TLR9 might therefore be more closely linked to Treg cell-related cytokine gene expression, but additional experiments are required in order to accept this hypothesis, including the analysis of the immune response against T. spiralis infection in TLR9-deficient mice. TLR9 is activated by unmethylated CpG dinucleotides that can be mimicked by synthetic oligonucleotides containing CpG motifs. CpG oligonucleotides (CpG ODNs) and CpG DNA can trigger intracellular signaling leading to the activation of macrophages, dendritic cells, and B cells, and the production of cytokines, chemokines, and immunoglobulins via TLR9 [7]. CpG ODNs have also been shown to stimulate the production of IL-10 by dendritic cells and promote the induction of Treg cells [17], and some parasite antigens can also initiate TLR9-dependent Treg cell activation [18-20]. T. spiralis has further been shown to induce Treg cell recruitment (with IL-10 and TGF-β as important cytokines) and regulate host Th2 and Th17 immune responses [4].
In conclusion, we observed that the gene expression levels of TLR4 and TLR9 are upregulated in the small intestine and muscle tissue of mice during T. spiralis infection. We suggest that TLR4 and TLR9 might be closely associated with Th2- and Treg cell-mediated immune responses, but additional data are needed for conclusive demonstration of this theory.