INTRODUCTION
Human sparganosis is a zoonotic disease caused by infection with the larval forms (procercoid/plerocercoid) of Spirometra spp. The first recorded case of human sparganosis in Korea involved a farmer [1] based on a report by Uemura in 1917. Cho et al. [2] reviewed 63 cases of indigenous sparganosis in 1975 in Korea. Then, an additional 56 cases were presented during the period from 1975 to 1989 [3]. The most recent report published in 2015 details human sparganosis cases recorded for the period 1979 and 2009 in Korea involved Spirometra erinaceieuropaei (Rudolphi, 1819) (n=35) and Spirometra decipiens (Diesing, 1850) (n=15) [4]. As a consequence, S. decipiens was added to the causative agent list of human sparganosis that was previously regarded as only involving S. erinaceieuropaei.
The routes of spargana infection involve either drinking water contaminated with procercoid-infected copepods or consumption of undercooked meat of plerocercoid-infected snakes or frogs. Additionally, infection can occur by placement of poultices that use the skin of infected snakes or frogs. Snakes have been noted as a very important source of infection for human sparganosis by many researchers in Korea [5-7]. At least 7 species of terrestrial snakes have thus far been investigated as possible sources of infections with spargana, among which the following 6 species of snakes have been positively identified with spargana: Elaphe rufodorsata, Elaphe dione, Dinodon rufozonatum ruforzonatum, Rhabdophis tigrinus tigrinus, Zamenis spinalis, and Agkistrodon halys. All of these snakes were from Gangwon-do (Province), which is located at the eastern side of the peninsula [8]. Spargana have been found in various species of frogs, snakes, birds, and mammals in the Far East region, including China and Japan, and 6 spirometrid species (under the name Diphyllobothrium) were reported with morphological and biological studies on the species in China [9]. The present study was to provide information for what Spirometra species are being distributed in Korea and China by using genetic analysis.
The morphological identification of spargana is usually difficult because they lack distinguishing characteristics. More recently, molecular approaches to differential identification of S. erinaceieuropaei and S. decipiens have been introduced that analyze sequence differences of mitochondrial genomes [4,10]. In the present study, we used species-specific primers for a multiplex PCR assay based on mitochondrial sequences of S. erinaceieuropaei and S. decipiens. The larval forms of Spirometra spp. (spargana) collected from terrestrial snakes in Korea and China were identified by the multiplex PCR assay and through phylogenetic analysis of mitochondrial DNA sequence data.
MATERIALS AND METHODS
Snakes
A total of 283 snakes were obtained from the provinces of Chungbuk, Chungnam, and Gyeongbuk in Korea and from unknown localities in China during the period from 1993 to 2001. Among these specimens, 161 Korean snakes (1993-2002) were donated by Association of Wild Animals Protection for research purpose and included R. tigrinus tigrinus (n=150), D. rufozonatum rufozonatum (n=4), Elaphe davidi (n=2), Elaphe schrenckii (n=2), and Agkistrodon saxatilis (n=3). The remaining 122 Chinese snakes had been smuggled from China and subsequently donated to Chungbuk National University School of Medicine by the Busan Customs of Korea in 2002 for research purpose and included D. rufozonatum rufozonatum (n=60), E. schrenckii (n=5), and A. saxatilis (n=57) (Table 1).
Nucleotide sequencing and multiplex PCR
A total of 904 plerocercoids were subjected to molecular biological analysis in this study. This included 50 plerocercoids for the analysis of mitochondrial cox1 gene sequences comprising 10 plerocercoids from each of 5 snake species. The remaining 854 plerocercoids were analyzed using a multiplex PCR assay. A single plerocercoid larva was chopped into small pieces on a glass slide, and total genomic DNA was extracted with a DNeasy tissue kit (Qiagen, Valencia, California, USA) according to the manufacturer’s instructions. PCR and DNA sequencing were performed according to protocols previously established [4].
For multiplex PCR, the forward primer, Se/Sd-7963F (5ʹ-ACG TGG TTT GTG GTG GCT CAT TTT-3ʹ, nucleotides 7963-7986 from specimen numbers KJ599679 and KJ599680 uploaded to GenBank) was designed from the highly conserved sequences of the S. decipiens and S. erinaceieuropaei cox1 gene. The first reverse primer Se-8344R, specific for S. erinaceieuropaei (5ʹ-TGG TCA CCT ATA CCC TAT CAT-3ʹ, nucleotides 8344-8364 from KJ599680) was designed to amplify different sized products from the second one. The second reverse primer was Sd-8584R, which was specific for S. decipiens (5ʹ-GTA TCA AGT TGG TTA GGA AGT TAA-3ʹ, nucleotides 8584-8607 from KJ599679). In order to test differential diagnosis of spirometrid tapeworms, multiplex PCR was performed using 15 DNA samples that comprised S. erinaceieuroapei plerocercoids (n=8) from humans and S. decipiens plerocercoids (n=7) from snakes. These DNA samples were confirmed by analysis of cox1 gene sequences.
DNA sequence analyses
DNA sequences were assembled and aligned with Geneious 6.1.5 (Biomatter, Auckland, New Zealand). The sequenced regions were identified by comparison with sequences of S. decipiens (KJ599679) and S. erinaceieuropaei (KJ599680), which were available in GenBank database. The phylogenetic analyses were performed with PAUP 4.0 [11] using the neighbor-joining (NJ), maximum parsimony (MP), and maximum likelihood (ML) methods, with D. nihonkaiense and D. latum as out-groups. Phylogenetic trees were constructed using the partial mitochondrial cox1 sequences (374 bp) of 5 taxa of Diphyllobothriidae as represented by S. decipiens (KJ599679), S. erinaceieuropaei (KJ599680), S. proliferum (AB015753), D. nihonkaiense (EF420138), and D. latum (DQ985706). Pairwise genetic distances based on the partial cox1 sequences (374 bp) were analyzed using the NJ method with the K2P substitution model. The parasite materials used in this study were provided by the Parasite Resource Bank of Korea, National Research Resource Center, the Republic of Korea.
RESULTS
Sparganum infection in snakes
The 283 snakes subjected to this study were identified as 1 of the 4 species of Colubridae; R. tigrinus tigrinus (n=150), D. rufozonatum rufozonatum (n=64), E. davidi (n=2), and E. schrenkii (n=7), or a species of Viperidae; A. saxatilis (n=60). With the exception of E. schrenkii and E. davidi, spargana were found in specimens of the remaining 3 species of snakes. The overall prevalence of spargana was 83% (235/283), with the highest infection rate being recorded for D. rufozonatum rufozonatum (100%), followed by A. saxatilis (85%) and R. tigrinus tigrinus (80%) (Table 1). A total of 904 plerocercoids were collected from these snakes. The average number of spargana per infected snake ranged 1.8-8.4, depending on the snake species.
Sequence divergence of mitochondrial cox1
The difference in nucleotide sequences of the partial cox1 gene between S. erinaceieuropaei (KJ599680) and S. decipiens (KJ599679) was 9.2%. Analysis of spargana specimens collected from snakes (n=50) revealed sequence identities with S. erinaceieuropaei and S. decipiens of 90.8% and 99.2%, respectively. The pairwise genetic distance between our spargana specimens (n=50) and S. decipiens ranged 0.0080-0.0107 while that with S. erinaceieuropaei ranged 0.1070-0.1096 (Table 2). Based on the phylogenetic relationships between the spargana, the Diphyllobothrium spp. were shown to be basal to the Spirometra spp. clades, as determined using the different analytical methods (MP, NJ, and ML) shown in Fig. 1. In the present study, all of the 50 spargana randomly chosen and comprised of 10 specimens from each of 5 species of snakes that included D. rufozonatum rufozonatum (Korean and Chinese), R. tigrinus tigrinus (Korean), and A. saxatilis (Korean and Chinese) were identified as S. decipiens using nucleotide sequence analysis of mitocondrial cox1.
Differential diagnosis of spirometrid tapeworms by multiplex PCR
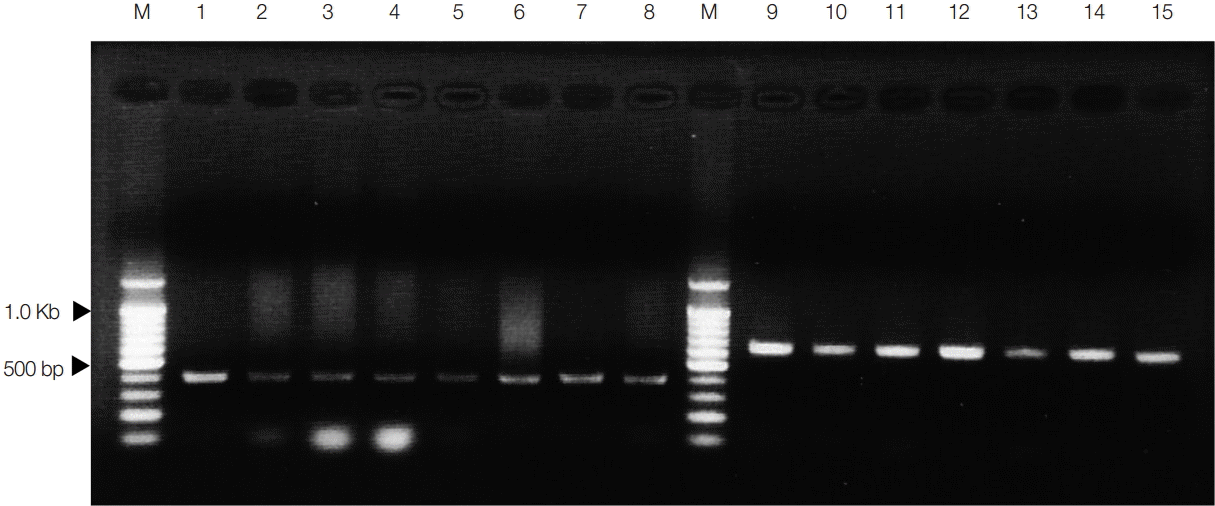
The results of PCR amplification of spirometrid tapeworm DNA using multiplex PCR are presented in Fig. 2. The PCR products obtained using the species-specific primers were positively detected in all PCR tested with a mixture of S. erinaceieuropaei and S. decipiens DNA templates. The S. erinaceieuropaeispecific bands (401 bp) were obtained in PCR with a mixture of S. erinaceieuropaei-specific primers (Se/Sd-7963F, forward; Se-8344bands, reverse) and S. erinaceieuropaei and S. decipiens DNA templates (lanes 1-8 in Fig. 2). S. decipiens-specific bands (644 bp) were also detected in PCR with a mixture of S. decipiens-specific primers (Se/Sd-7963F, forward; Sd-8584R, reverse) and S. decipiens and S. erinaceieuropaei DNA templates (lanes 9-15 in Fig. 2). These PCR products were cloned and sequenced to confirm the specificity of each species-specific primer. The results of sequence analyses for the species-specific bands revealed a 100% sequence identity with homologous regions of the mtDNA sequences of S. erinaceieuropaei and S. decipiens. In order to identify spargana species, multiplex PCR was applied to the remaining 854 spargana. The species-specific 644-bp diagnostic band was detected in all of the 854 spargana subjected to multiplex PCR (data not shown).
DISCUSSION
It is known that the majority of human sparganosis cases are caused by consumption of raw reptiles (snakes), amphibians (frogs), or drinking water contaminated with larval Spirometra species. Regarding the well-known species Spirometra erinaceieuropaei, Cho et al. [2] investigated 56 cases in search of possible sources of infections for human sparganosis. Approximately a half of the cases (n=30) were involved with consumption of raw snakes, and 10 cases were associated with drinking untreated water. Thus, the consumption of raw snake flesh has been considered the most important cause of human sparganosis in Korea. This was generally accepted by most parasitologists without opposition; however, our research group recently suggested that S. erinaceieuropaei is not the only species that induces human sparganosis [4]. The suggestion was that S. decipiens is another possible cause of human sparganosis in Korea. The species ratio presented in the report was 35 (S. erinaceieuropaei): 15 (S. decipiens) for cases of human sparganosis among Korean patients recorded for the period 1979-2009 from all of the parasitology laboratories in Korea. The simplified ratio of 2.3:1 reveals approximately twice as many cases of S. erinaceieuropaei than S. decipiens among the 50 specimens examined [4]. This finding immediately provoked questions concerning whether S. erinaceieuropaei and S. decipiens are caused by the same source of infection, namely, snakes, and if so, what is the ratio between these Spirometra species in the snakes? To answer these questions, genetic analyses of spargana that infect snakes are required to identify the Spirometra species. Our present study revealed a rather surprising result that all spargana from snakes were identified as S. decipiens (n=904) without exception. This finding highlights a serious epidemiological discrepancy between the ratios of Spirometra species in humans and snakes.
We do not currently know the reason why we did not find any S. erinaceieuropaei from any of our snakes. One possible explanation is that the S. erinaceieuropaei larval stage may be the primary source of human infection by way of drinking water contaminated with procercoids, but this then begs an even bigger question: what is the intermediate host that transfers larval forms to humans if it is not snakes? Furthermore, what is the final host? However, all these questions are based on a supposition that there are 2 different kinds of Spirometra in humans. The use of a poultice, which represents another form of spargana transmission to humans, was not taken seriously because it seems that it is not commonly practiced nowadays. The final hosts of Spirometra spp. are commonly known to be carnivorous animals such as cats and dogs, but S. erinaceieuropaei has not been found or ever described in natural infections in cats and dogs [9]. Thus, S. decipiens might be the only species of naturally occurring tapeworm that is currently found in cats and dogs. The only clearly known second intermediate host of S. erinaceieuropaei is the Chinese hedgehog. These findings indicate that further studies are required to determine the real infection route and final hosts of S. erinaceieuropaei, and to clarify whether S. decipiens infects humans through drinking water contaminated with procercoids.
In the present study, the differential diagnosis obtained by PCR using species-specific primers was correlated with species identification based on nucleotide sequence analysis of the mitochondrial cox1 gene. The results showed that the multiplex PCR assay with the Se/Sd-7963F, Se-8344R, and Sd-8584R primers will be useful for species identification in the genus Spirometra. Finally, the spargana that infected snakes (D. rufozonatum rufozonatum, R. tigrinus tigrinus, and A. saxatilis) in Korea and China was S. decipiens, and this species was distinguished from the spargana of S. erinaceieuropaei using analysis of mitochondrial cox1 sequences and multiplex PCR. We wonder if our mysterious finding that S. erinaceieuropaei larval forms were not identified in snake specimens collected from Korea and China will be explained in the near future if investigations use larger sample sizes.