INTRODUCTION
Trichinellosis caused by Trichinella spiralis is a globally distributed disease. There have been reports of outbreaks from China, Japan, Thailand, Laos, Vietnam, Korea, Eastern Europe, and West Africa [1,2]. Human infection occurs through ingesting T. spiralis larvae that are encysted in muscle tissue of meat from domestic or wild animals. Although reinfection with T. spiralis occurs frequently in domestic and wild animals with occasional human T. spiralis outbreak, the host responses to reinfection with T. spiralis remains largely unknown [1,3,4].
Antibody responses against T. spiralis in rats are dose-dependent, showing a positive correlation between infection dose and serum antibody levels [5]. In hosts, such as cattle, sheep, goat, horses, wild boars, pigs, and foxes, antibody responses induced by T. spiralis infections have also been documented to be dose-dependent [6–16]. A study investigating the resistance against reinfection reported the reductions of T. spiralis in the small intestine and muscle for adults and larvae, respectively. In this study, only a single high dose of T. spiralis larvae (400 larvae per mouse) were used for primary and secondary infections in BALB/c mice [17]. Since humans or animals could be infected more frequently by a low number of larvae in natural conditions [18], the study of host responses induced by different dose of infection, by a low dose of T. spiralis infection in particular, would have an important impact.
A recent study indicates that eosinophils served to limit the number of new born larvae of T. spiralis that migrated in tissue and colonized in skeletal muscles [19]. Eosinophils are required for control of a secondary infection by T. spiralis, and the effect is dependent on the presence of specific antibodies [19]. Thus, it would be of utmost importance to clarify the host resistance to reinfection, including host eosinophil responses with T. spiralis-specific antibody responses. Circulating CD23+ B cell subset correlates with the development of resistance to Schistosoma mansoni reinfection [20]. Memory B cells reside in secondary lymphoid organs, where they can rapidly mobilized upon a new antigenic encounter. Memory B cells are composed of 2 subsets, an IgM+ and an IgG+ subset. The IgG subset comprises of IgG1 cells, which represent the major fraction of antigen-specific switched memory B cells. However, the existence of a long-lived IgM+ memory subset, in addition to IgG+ memory set, was found in a mouse model recently [21]. Thus, a study on CD23+ B cell and IgM+ memory subset-related resistance against T. spiralis infection along with antibody response and eosinophil response would have a significant impact.
MATERIALS AND METHODS
Animal and parasite
Female inbred BALB/c (aged 8 weeks; Nara Biotech, Kyonggi-do, Korea) were used. Groups of mice (n=10) were orally infected with 10 larvae (10 TS) or 100 larvae of T. spiralis (100 TS). The parasites used in this study were from T. spiralis Korea isolate [22] and maintained by serial passage in SD rats. Muscle tissues were collected and muscle stage larvae were isolated by artificial digestion by using pepsin-HCl at 45±1°C.
Primary infection and challenge infection
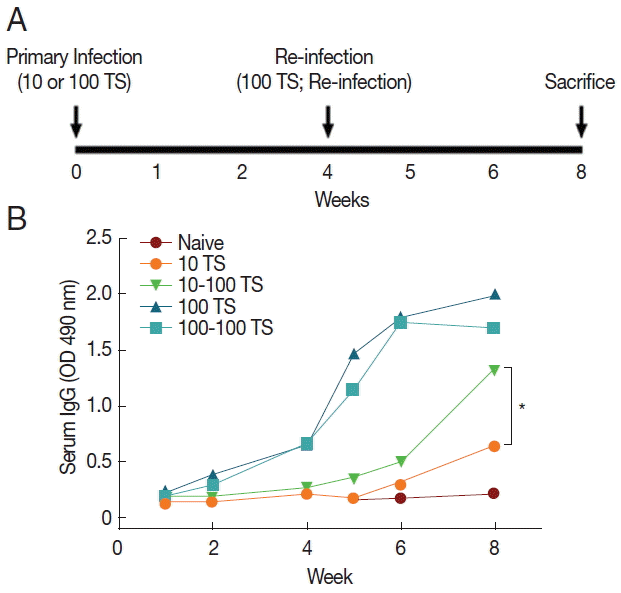
Groups of mice (n=10) received primary infection with 10 TS or 100 TS via an oro-gastric tube. Challenge infections (reinfection) were performed at week 4 after primary infections as indicated in Fig. 1A. At week 8, mice were sacrificed, and the diaphragms were collected. The diaphragm from individual mouse was digested with pepsin-HCl at 37°C overnight, and T. spiralis larvae were harvested and counted. All animal experiments and husbandry involved in these studies were conducted under the guidelines of the Kyung Hee University IACUC, Seoul, Korea. Kyung Hee IACUC operates under the National Veterinary Research and Quarantine Service (NVRQS) and regulations of the World Organization for Animal Health (WOAH).
T. spiralis excretory/secretory antigen preparation
T. spiralis excretory/secretory (ES) product was produced as described previously [23,24]. T. spiralis larvae were isolated from mouse muscle tissue by artificial digestion. Clean larvae were incubated in a CO2 incubator for 24 hr at 37°C in a petri dish containing RPMI-1640 culture medium without fetal bovine serum (FBS). The culture supernatants were collected by centrifugation. The supernatant was dialyzed and lyophilized. The protein concentration was determined by DC Protein Assay Kit (Bio-Rad, Hercules, California, USA), and T. spiralis ES products were stored at −70°C until use.
Worm burden determination
Mice were sacrificed at day 7 and week 4 after challenge infection. Intestine from individual mice was collected and cut into 10 cm long pieces, placed into dish, and incubated in 0.9% saline at 37°C for 2 hr. After incubation, intestines were discarded, and the sediment was counted under stereomicroscopy. The mouse diaphragm from individual mouse was collected and digested with pepsin-HCl at 37°C overnight. T. spiralis adults and larvae were harvested and counted under a microscope.
Antibody responses
Blood samples were collected by retro-orbital plexus puncture at week 1, 2, and 4 after primary infection and at week 1, 2, and 4 post-challenge infections as indicated in Fig. 1A. T. spiralis-specific serum IgG antibodies were determined by ELISA, as described previously [25]. Briefly, 96-well microtiter plates (Nunc Life Technologies, Basel, Switzerland) were coated with 100 μl of ES product at a concentration of 4 μg/ml in coating buffer (0.1 M sodium carbonate, pH 9.5) at 4°C overnight. The plates were then incubated with mouse sera (1:100) from infected mice. Horse-radish peroxidase-labeled goat anti-mouse IgG (1:2,000) (Zymed, San Francisco, California, USA) was placed into plates and incubated at 37°C for 1.5 hr. Finally, the substrate o-phenylenediamine (Zymed) in citrate-phosphate buffer (pH 5.0) containing 0.03% H2O2 (Sigma) was used for color development. The optical density at 490 nm was read using an ELISA reader (Molecular Devices, Sunnyvale, California, USA).
Detection of antibody-secreting cells (ASCs) in the spleen
Freshly isolated spleen cells (1×106 cells) were added to a 96-well culture plate and incubated for 3 days at 37°C under 5% CO2 to detect T. spiralis-specific ASCs. ASCs were detected in the supernatant using an ELISA plate coated with T. spiralis ES antigen. Horseradish peroxidase (HRP)-conjugated secondary goat-anti-mouse antibodies were added and incubated. The substrate o-phenylenediamine (Zymed) was used as a measure of HRP activity, and optical density was read at 490 nm.
Flow cytometry analysis
At week 4 post-challenge infection, mice were euthanized to collect spleen cell samples. For cell phenotype analysis, cells from spleen (n=5) were stained with fluorophore-labeled surface markers. Anti-mouse CD16/32 (clone 2.4G2, BD Pharmingen, San Diego, California, USA) was used as Fc receptor blocker. Afterwards, antibody cocktails which contained anti-mouse CD23-FITC (clone 53-6.7, BD Pharmingen) and anti-mouse IgM-PerCP-Cy™5.5 (clone R6-60.2, BD Pharmingen) were used to treat cells. The stained cells washed after incubation, acquired by flow cytometer LSRFortessa (BD Biosciences, San Diego, California, USA) and analyzed using FlowJo program (Tree Star Inc., San Carlos, California, USA).
Eosinophil counts
Whole blood was collected from mice at week 1, 2, and 4 after primary infection and week 1, 2, and 4 after challenge infection. Whole blood was stained with Discombe’s solution. A 20-μl of individual blood was immediately added into 180 μl of Discombe’s solution (5 vol. acetone, 5 vol. 1% aqueous eosin solution, 90 vol. distill water) and mixed well. Eosinophils in whole blood were counted under microscopy using a hemocytometer.
RESULTS
Antibody responses in immune serum
The time points for primary infection, reinfection, and sacrifice were seen in experimental schedules (Fig. 1A). Mouse sera collected at different time points were used to measure the levels of T. spiralis-specific antibody responses. The levels of total IgG in the serum specific T. spiralis ES Ag after primary infection and reinfection were determined (Fig. 1B). Total IgG antibody responses from mice primarily infected with 10 T. spiralis (10 TS) showed significantly higher titers after reinfection (10–100 TS) compared to those after primary infection (10 TS) (*P<0.05, Fig. 1B). With higher dose of primary infections (100 TS), no significant difference was found (Fig. 1B). These results indicated that mice primarily infected with 10 TS induced significantly higher levels of T. spiralis-specific antibodies upon reinfection.
Antibody secreting cell responses
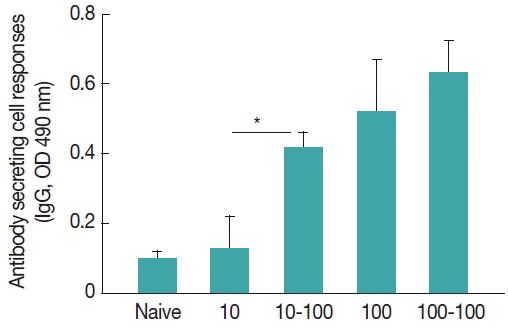
During B cell differentiation and development, a fraction of germinal center B cells traffics to the bone marrow or spleen and develops into antibody secreting plasma cells (APC). To investigate the induction of antibody secreting cells, spleen cells were harvested at week 4 after challenge infection and stimulated in vitro with T. spiralis ES Ag (Fig. 2). Mice with 10–100 TS showed higher levels of IgG antibodies bound to the T. spiralis ES Ag (*P<0.05). Overall, low dose infection (10 TS) induced increased antibody secreting cell responses after reinfection, consistent with the results of IgG antibody responses in sera.
Resistance against reinfection
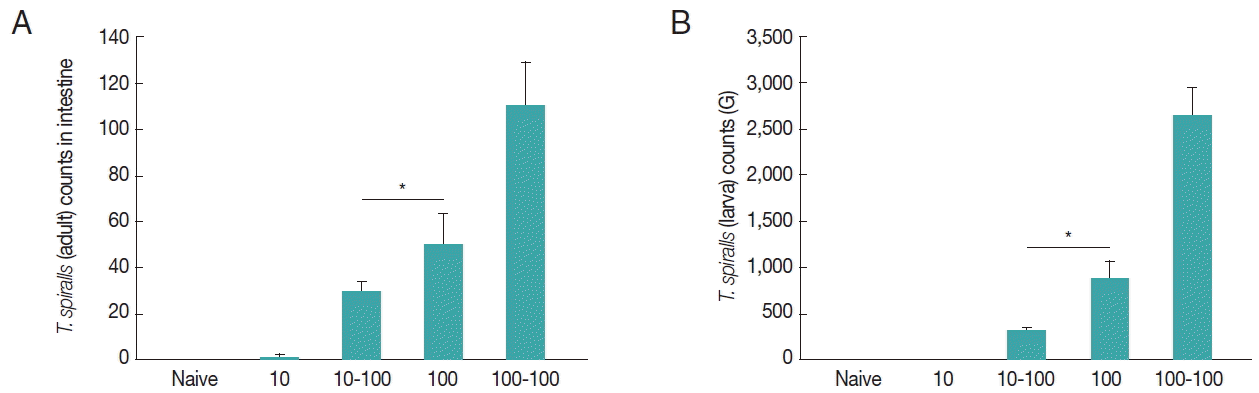
The worm burden following challenge infection is the most important indicator to assess protective immunity. Mice were reinfected with 100 larvae of T. spiralis at week 4 after primary infection and worm burdens at day 7 and week 4 post-challenge were determined. As shown in Fig. 3A and 3B, significantly decreased worm burdens were detected in mice primarily infected with 10 TS compared to primary controls (Fig. 3A, B, *P<0.05). However, no significant worm burden decrease was detected in mice primarily infected with 100 TS. These results indicated that mice primarily infected with 10 TS can effectively inhibit T. spiralis replication, whereas mice primarily infected with 100 TS cannot inhibit T. spiralis replication, indicating that different does of T. spiralis in mice induces different levels of resistance.
B cell responses
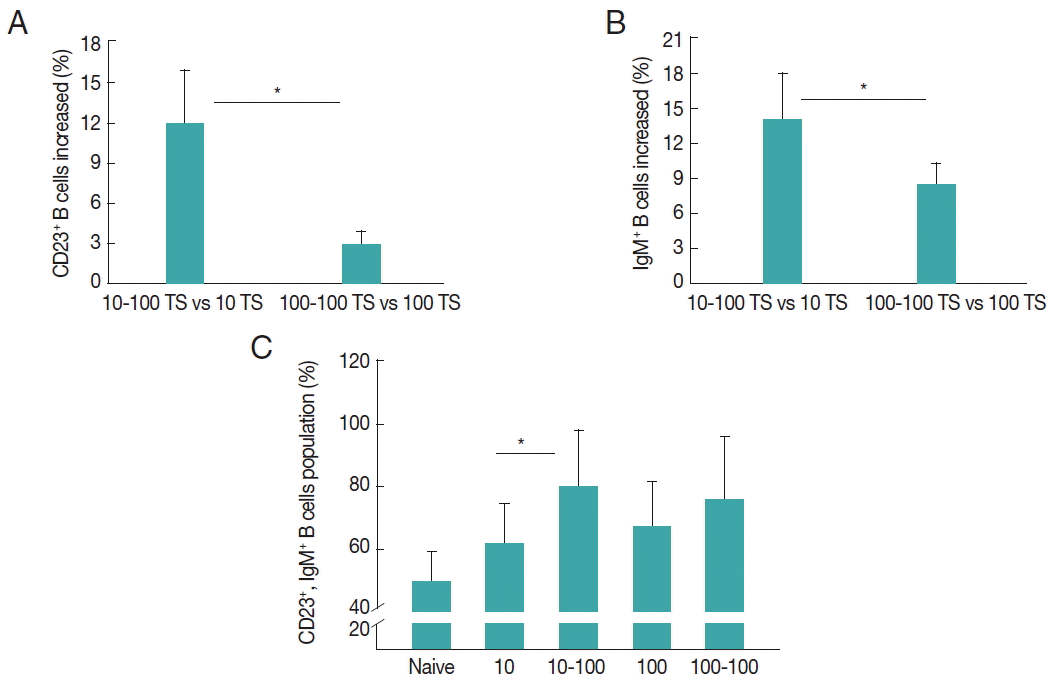
The populations of CD23+ B cells and IgM+ B cells in spleen were measured as shown in Fig. 4. Mice were primarily infected with 10 TS or 100 TS, then challenge infected with 100 TS. Upon challenge infection, mice with a low dose of infection (10 TS) showed significant increases of CD23+ and IgM+ B cells, respectively, compared to mice with 100 TS infection (*P<0.05, Fig. 4A, B). Importantly, a higher level of double positive CD23+IgM+ B cells was only seen in mice with 10–100 TS infection, not in mice with 100-100 TS (Fig. 4C), indicating double positive CD23+IgM+ B cells might contribute the resistance induced by a light infection.
Eosinophil profiles after primary infection and reinfection
Whole bloods were collected as indicated in methods section and stained with Discombe’s solution. As shown in Fig. 5, eosinophils raised significantly at week 2 after primary infection with both 10 TS and 100 TS (Fig. 5A, B, *P<0.01). Slightly increased eosinophil level was observed at week 4, which subsequently decreased in the following weeks. Upon challenge infection, eosinophils levels rose again at week 1 after challenge, reached its highest levels at week 2, and then decreased at week 4 (Fig. 5A, B). Nonetheless, the challenge infection eosinophil count far exceeded those of primary infection eosinophil count at week 6 in 10 TS mice (Fig. 5A; ***P<0.05). Interestingly, at week 8, 10 TS showed significantly higher numbers of eosinophil counts upon reinfection, whereas no significant increase of eosinophil was detected in the case of 100 TS mice (Fig. 5A, B; **P<0.01), indicating that the eosinophils from 10 TS may contribute to the resistance induced by 10 TS primary infection. The results indicated that significantly increased eosinophils at weeks 2 and 4 post-reinfection from 10 TS primary infection are protective.
DISCUSSION
In this study, we assessed the resistance of mice against T. spiralis reinfection by determining the larval worm burden reductions in the intestines for adults and in the diaphragms for larvae. Our results indicated that a dose of 10 TS infection induced significant resistance against reinfection, which correlated with IgG antibody response, CD23+ B cells, IgM+ B cells, and eosinophil responses. To our knowledge, this is the first report to identify the immune correlates of resistance to T. spiralis reinfection induced by a low dose of T. spiralis primary infection.
The dose of T. spiralis larvae used for the primary infection seems critical for developing parasitic resistance. In our study, only mice primarily infected with 10 TS showed enhanced T. spiralis-specific IgG antibody responses upon challenge. Because antibody response is dose-dependent as described by previous studies [5], observing a higher serum IgG level from 100 TS primary infection than 10 TS was reasonable. Yet, challenge infection with 10 TS mice with 100 TS increased the serum IgG level nearly 2-fold by week 8, whereas no significant enhancement was observed in 100-100 TS mice (Fig. 1B). A similar pattern was observed for antibody secreting cell responses as well, and combining these results, it can be assumed that these antibody responses may have led the host to acquire parasitic resistance.
Repeated exposures and treatment of Schistosoma mansoni have been documented to eventually lead to higher levels of resistance towards the parasite [26], and the same can be applicable to our study. With dramatic worm burden difference between mice with 10–100 TS and 100 TS (Fig. 3A, B), it is highly plausible that a previous exposure to 10 TS have resulted in a greater resistance to reinfection. Results from our study showed that significantly high levels of eosinophils were detected at week 2 and 4 after primary infection and reinfection. At week 4 post-reinfection, mice primarily infected with a low dose of T. spiralis (10 TS) showed a higher number of eosinophils compared to those with 100 TS (Fig. 5A, B), indicating that eosinophils are required in developing resistance towards T. spiralis reinfection [19].
In addition to eosinophils, we found that the resistance to reinfection correlated with elevated T. spiralis-specific antibody response, CD23+ B cells and IgM+ B cells responses, indicating eosinophils are involved in the resistance to reinfection by binding themselves to larvae in the presences of antibodies [19]. Humoral memory is maintained by memory B cells and plasma cells. Reinfection triggers the formation of antibody-forming cells (AFC) [21]. In our study, significantly higher levels of antibody secreting cells were found in mice with 10–100 TS (Fig. 2), indicating memory B cells differentiated into plasma cells and produced antibodies upon reinfection. CD23+ B cells are reported to be correlated with the development of resistance to S. mansoni reinfection and eosinophils [20], which is consistent with our results. IgG1 memory B cell has been taken as a surrogate marker for memory in the mouse [21]. However, a recent study revealed the existence of a long-lived IgM memory B cell population in mouse in addition to classical IgG+ cells [21]. This study is consistent with our results as significantly raised IgM+ B cell response was observed upon reinfection, which may indicate that IgM+ B cells might contribute to resistance.
Memory B cells and parasite-specific antibody are critical for development of parasitic resistance. A study on Trichuris muris has shown that B cells are a requirement for parasite resistance [27], and mitigating parasitic invasion through antigen-specific antibody towards T. spiralis has been confirmed in vitro [28,29]. Furthermore, parasite-specific antibodies and its associated Th2 responses limit worm establishment and may even play a role in diminishing the effect of challenge infections [27]. Similarly, splenic antibody response towards Haemonchus contortus reveals that resistance mounted towards the parasite in sheep is reliant on Th2 type immune response [30]. However, these previous studies largely focused on primary infections, and the correlations of immune response profiles with the resistance against reinfection have not been clarified.
In conclusion, our study has provided new insight into resistance induced by a low dose (10 TS) of T. spiralis. In the present study, we have demonstrated that upon reinfection, mice primarily infected with 10 TS induced enhanced IgG antibody, CD23+ B cells, IgM+ B cells, and eosinophil responses, resulting in significant reduction of T. spiralis burden, thereby indicating that all of these factors are correlated with the resistance against reinfection by T. spiralis.