INTRODUCTION
Malaria is an infectious disease affecting humans, which is transmitted by the bite of Anopheles mosquitoes harboring sporozoites of parasitic protozoans belonging to the genus Plasmodium. Five species, including Plasmodium falciparum, P. vivax, P. ovale, P. malariae, and P. knowlesi, infect humans in the respective endemic areas [1,2]. The parasitic disease remains a significant health threat with more than 3 billion people at risk of infection per year. However, substantial progress has been made in fighting malaria and thus, according to the latest estimates between 2000 and 2015, case incidence and mortality rate of the protozoan disease were reduced by 41% and 62%, respectively. During the period, the numbers of country, where malaria is endemic, was also decreased from 108 to 91 countries. In 2015, 212 million cases occurred globally leading to 429,000 deaths, most of which were children aged under 5 years in Africa [3].
Despites the obvious progress in antimalarial interventions, emergence and spread of drug-resistant parasites, as well as insecticide-resistant mosquitoes, are still existing risks that potentially threaten the recent gains in malaria control [4]. Approaches to develop molecules that effectively protect transmission of malaria from mosquito vector and/or generate defensive immune response in humans have been recognized as the greatest challenges in malaria elimination [5–7]. By virtues of these foresight activities, numerous Plasmodium proteins have been targeted for the development therapeutic vaccines by blocking any of the vector- or host-specific malarial stages. Some of the vaccine candidates have already entered diverse phases for clinical testing [3,7].
Of the 5 Plasmodium species infecting humans, P. falciparum is the most significant cause of malaria and thus has become the major concern in the relevant areas investigating molecular tools for malaria control. However, studies on human malaria are inherently restricted by difficulty either in using humans as an experimental system or in maintaining in vitro the complete life cycle of the protozoan parasites. Rodent malaria parasites, including Plamodium berghei, P. yoelii, P. chabaudi, and P. vinckei can be a practicable choice to use as models of human malaria in the related experiments [8,9]. Small but significant biological difference among them in laboratory mice makes these rodent malaria particularly attractive models for the investigation of specialized aspects of human malaria. For example, P. berghei preferentially invades reticulocytes and induces severe pathology in mice [8]. P. chabaudi can be an appropriate model in studies associated with the drug resistance and immune evasion especially by antigenic variation [10]. In contrast, P. yoelii has been widely used in experiments concentrating on the liver-stage biology of malaria and on the host immune responses against that stage [9,11,12]. The last species is also highly applicable either in the investigations of receptors involved in erythrocyte binding [13] or in the evaluation of vaccine candidates [14–16].
In this study, we identified and cloned P. yoelii genes, including merozoite surface protein 1 (MSP1), circumsporozoite protein (CSP), and var 2 chondroitin sulphate A (VAR2CSA) genes, which can be applied in the investigation of immunization-induced protective immunity against P. yoelii malaria. The presence of antibodies specific to the recombinant fragments of MSP1 and CSP proteins was also examined in mouse sera infected with P. yoelii.
MATERIALS AND METHODS
Experimental infections
Male ICR mice (4-week-old) were housed in the experimental animal center of Ewha Womans University School of Medicine, Seoul, Korea under the Institutional Animal Care and Usage Committee (IACUC) approved protocol for the animal care. Parasitemia of the non-lethal 17XNL strain of P. yoelii (courtesy of Prof. ET Han, Kangwon National University, Chuncheon, Korea) was monitored by microscopic examination of Diff-Quik stained blood smears from passage mice and expressed as the percentage of parasitized RBCs. Blood-stage infection was initiated by intraperitoneal injection of 150 μl of the parasitized erythrocytes (5.5%). From day 2 post challenge, the resulting parasitemia was periodically monitored by enumerating parasitized erythrocytes in thin tail-blood smears stained with Giemsa, as previously described [17]. A total of 6 experimental mice that were recovered to normal levels 3 weeks after the primary injection were reinfected with the P. yoelii cells (5.5%) at 4 weeks after the first infection. Changes in the ear color and parasitemia were similarly checked in the experimental group.
Parasite materials
Blood was collected through cardiac puncture from experimental mice (n=6) at day 10 after the primary and secondary infections of P. yoelii 17XNL strain. Plasma samples prepared from the blood were used as the 1st and 2nd antisera, respectively, specific to P. yoelii antigens. Blood was also collected from uninfected normal mice with the same age (n=6) to prepare the normal control sera. The sera were stored at −80°C until use.
The trophozoite and schizont stage parasites in infected mice blood from the primary and secondary infections (parasitemia >35%) were highly enriched by the Optiprep (iodixanol; Axis-shield, Oslo, Norway) density gradient, as previously described [18]. Cellular fraction of normal mice blood was also prepared to use as control mouse proteins. Proteins were extracted by incubating the cellular fraction samples in a protein lysis buffer (155 mM NH4Cl, 12 mM NaHCO3 and 0.1 mM EDTA) containing 1% SDS and protease inhibitors (Roche, Mannheim, Germany) for 1 hr on ice. The solutions were centrifuged at 12,000 rpm for 10 min at 4°C and the centrifugal supernatants were taken as protein samples. The total protein concentrations were determined by the Lowry method and stored at −80°C until use.
Genomic DNA of P. yoelii 17XNL was extracted from whole blood collected from infected mice (parasitemia>35%) using the QIAamp DNA Blood Mini Kit according to the manufacturer’s instruction (Qiagen, Hilden, Germany). The concentration and quality of the prepared genomic DNA was determined by a spectrophotometer by measuring absorbance at 260 and 280 nm. The DNA sample was stored at 4°C until use.
Identification of P. yoelii genes
As vaccine candidates applicable in the examination of protective immunity, 4 malaria proteins (MSP1, CSP, VAR2CSA, and liver stage antigen 1 [LSA1]) were selected in this study. P. yoelii genes homologous to the selected molecules were searched in the genomic database of the rodent malaria in PlasmoDB (http://plasmodb.org/plasmo/) using BLAST algorithms. Protein and mRNA sequences used as queries in the homology searches were retrieved from the GenBank (https://www.ncbi.nlm.nih.gov/) database of Plasmodium spp. (no. AAA66185.1 for MSP1, partial sequence of P. yoelii protein; no. AAY34563.1 for CSP, P. yoelii; no. ALB01043.1 for VAR2CSA, P. falciparum; and no. CD064792.1 for LSA1, P. reichenowi). The genomic nucleotide sequences were compared to the respective mRNA sequences and then, their genomic structures were determined by referencing the general GT-AG rule [19].
Amplification of full-unit genes
Forward and reverse primers were designed to match the 5′- and 3′-ends of P. yoelii genes. The primer sequences were as followings: 5′-ATGAAGAAGTGTACCATTTTAGTTGTAGCGTC-3′ and 5′-TTAATTAAAGAATACTAATACTAATAATATTAC-3′ for CSP gene; 5′-ATGAAGGTGATTGGACTTTTATTTTCTTTCG-3′ and 5′-TTAAAATATATTAAATACAATTAATGTGATAA-3′ for MSP1 gene; 5′-ATGAACAAAATAATAACTTATTTTTCTTTTATAC-3′ and 5′-TTATTTTAAAAATATGACTATGCATACCATC-3′ for VAR2CSA gene. The primers were used in long-range PCR to amplify each of the full-length genes from the genomic DNA of P. yoelii. The typical 2-step thermal cycling PCR was performed for the amplification using the LA Taq polymerase (Takara, Shiga, Japan) (initial denaturation at 95°C for 5 min, 35 cycles of 45 s at 94°C and 5 min at 64°C, and a final extension at 70°C for 10 min). After verifying their correct sizes and purity on 0.8–1%-agarose gels, the products were cloned into a pGEM-T Easy vector (Promega, Madison, Wisconsin, USA). The nucleotide sequences were automatically determined using a BigDye Terminators Cycle Sequencing Core Kit (Ver. 3·0; Perkin Elmer, Foster City, California, USA) and an automated ABI PRISM 377A DNA Sequencer (Applied Biosystems, Foster City, California, USA).
Sequence analysis
Open reading frame (ORF) encoded in each of the cloned genes was predicted using ORF Finder (http://www.ncbi.nlm.nih.gov/) and their similarity profiles were further verified using BLASTp with the theoretically translated amino acid (aa) sequences. The aa sequences of orthologous proteins identified in the PlasmoDB and GenBank databases were aligned with ClustalX and used in phylogenetic analysis with MEGA 6.0 (neighbor-joining algorithm, Jones-Thornton-Tayler model for aa substitution, rate heterogeneity with 4 gamma category, pairwise-deletion of gapped region sequences). The statistical reliability of branching nodes was assessed by the standard bootstrapping test of 1,000 replicates. The aa sequences were also applied in the prediction of functional protein domains using the SMART program (http://smart.embl.de/).
Generation of recombinantMSP1 and CSP proteins
The coding DNA sequences of MSP1 and CSP genes were segmentally amplified from the genomic DNA of P. yoelii with specific primers containing restriction sites for endonucleases (Supplementary Table 1). After digestion with the corresponding enzymes, the amplicons were cloned into pET-28a or pGEX-6p-1 plasmid and transformed into competent Escherichia coli BL21 (DE3) cells. Expression of these recombinant proteins were induced by incubating the cells in Luria-Bertani medium supplemented with 0.5 mM isopropyl-β-D-thiogalactopyranoside (IPTG) for 4 hr at 37°C. The proteins were purified by nickel-nitrilotriacetic acid (Qiagen) or glutathione sepharose (GE Healthcare, Little Chalfont, UK) chromatography according to the manufacturers’ instructions.
Western blot analysis
The cellular proteins (each 20 μg) extracted from the bloods of normal and infected mice were resolved through 10% SDS-PAGE, together with a pooled normal mice serum (n=5). The protein bands were either visualized by staining with Coomassie brilliant blue R-250 or transferred electrophoretically onto nitrocellulose membranes. The membranes were reacted with the specific mouse antisera prepared during the primary and secondary infections of P. yoelii 17XNL, as well as normal mouse serum (1:1,000 dilution). Antigen-specific antibodies were detected with the goat anti-mouse IgG antibody conjugated with IR-Dye 800cw (1:20,000 dilution). The positive reaction was scanned on an Odyssey CLx imager (800 channel; Lincoln, Nebraska, USA). Images were acquired on the CLx imager at 169 μm resolution. The recombinant MSP1 and CSP fragments (each 200 ng) were similarly applied in the western blot analysis.
RESULTS
P. yoelii MSP1 gene (PyMSP1)
A partial protein sequence (459 aa, 2104291A) corresponded to the N-terminal portion of PyMSP1 was identified in the GenBank database [20]. The sequence was used as a query during the tBLASTn analysis of P. yoelii transcripts in PlasmoDB to isolate the full PyMSP1 gene sequence. A total of two 5,319-bp transcripts (sequence identity=100%) were detected in the similarity analysis (PY05748 of P. yoelii yoelii 17XNL and PYYM_0834100 of P. yoelii yoelii YM). By using the sequence as a query in BLASTn search, the full-length sequence of a single exonic PyMSP1 gene was identified in the genomic databases of P. yoelii 17XNL/17X and P. yoelii YM strains (sequence identity=100%). The full gene sequence was amplified by a long-range PCR from the genomic DNA of P. yoelii 17XNL (Supplementary Fig. 1).
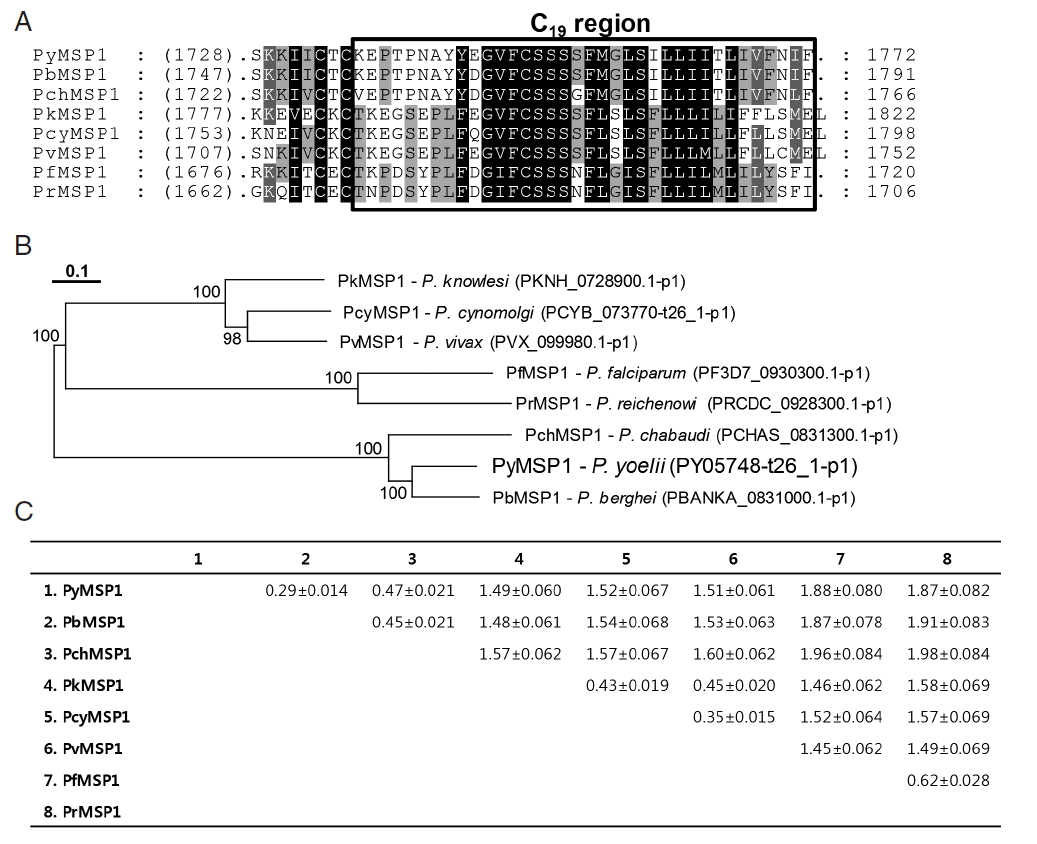
The 1772-aa PyMSP1 protein detected orthologous proteins in almost all of the Plasmodium databases in PlasmoDB, except for that of an avian malaria P. gallinaceum, and exhibited identity values ranging from 32% (P. falciparum MSP1) to 74% (P. berghei MSP1) with its plasmodial orthologs. As previously shown during characterization of P. berghei MSP1 [21], PyMSP1 also contained the structural/functional domains conserved in the plasmodial MSP1 proteins (data not shown). In a phylogenetic analysis, plasmodial MSP1 proteins showed a clustering pattern identical to that of their donor organisms [22]. The distance values were similarly estimated as 0.403± 0.1028 and 0.408±0.5110 among MSP1 proteins identified in rodent and primate malaria, respectively. However, the value was greatly increased to 1.535±0.0398 between rodent and primate malaria proteins (Fig. 1).
P. yoelii CSP gene (PyCSP)
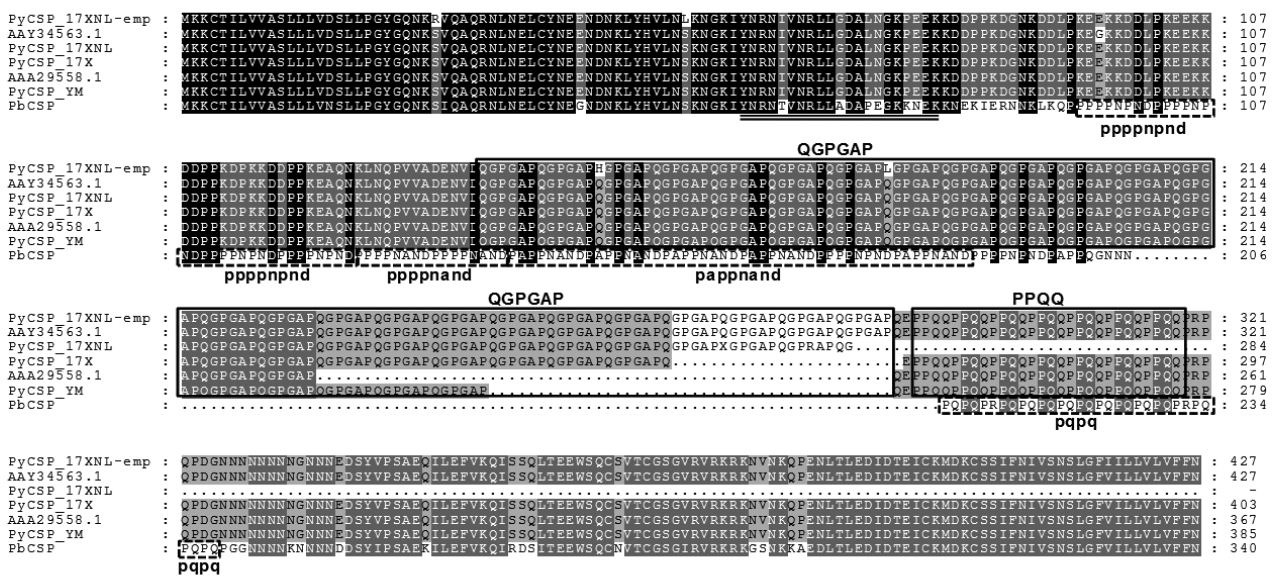
The genomic sequence of PyCSP gene was searched in PlasmoDB using BLAST programs with the mRNA (1284 bp, DQ012939.1) and aa (427 aa, AAY34563.1) sequences of P. yoelii CSP, which had been isolated from GenBank. The gene was also found to be single exonic. However, in contrast to MSP1, there were several gaps with various lengths in the resulting sequence alignment between the mRNA query and matched genomic sequences. Gap lengths in the alignment were slightly different among genomic sequences of P. yoelii strains, mainly due to the nucleotide length polymorphism in the repetitive region detected in the middle region of rodent malaria CSP genes [23]. The full-unit PyCSP gene empirically determined (PyCSP_17XNL-emp; Supplementary Fig. 1), had a size identical to that of DQ012939.1, although 7 nucleobase mismatches were observed between them (99.5% identity). Of the 7 mismatched bases, 5 bases changed the decoding amino acids (see PyCSP_17XNL-emp and AAY34563.1 in Fig. 2).
To verify the length polymorphism among the PyCSP proteins of different P. yoelii strains, orthologous sequences were isolated from PlasmoDB and GenBank. Two unique sequences were identified in 17XNL (PY07368-t26_1-p1/XP_728216.1 and AAY34563.1) and 17X (PY17X_0405400.1-p1/CDZ 10959. 1 and AAA29558.1) strains, whereas only a single unique sequence was detected in the proteomic database of YM strain (PYYM_0405600.1-p1/CDU16435.1). Sequence alignment of these PyCSP proteins together with that of P. berghei (PbCSP) demonstrated that the length polymorphism was induced by shrinkage/expansion of the repeat unit ‘QGPGAP’ found in the middle region of PyCSP (Fig. 2). It was also apparent that the repeat-region sequences, but not the 5′- and 3′-flanking regions, has been specifically evolved in each of the rodent malaria species. CSP proteins identified in primate malaria also exhibited repeat sequences specific to the lineages (data not shown). Amino acid sequence acting as the cytotoxic T lymphocyte epitope was tightly conserved in the PyCSP proteins with different strain origins (double underlined in Fig. 2) [24].
VAR2CSA and LSA1 genes in Plasmodium spp
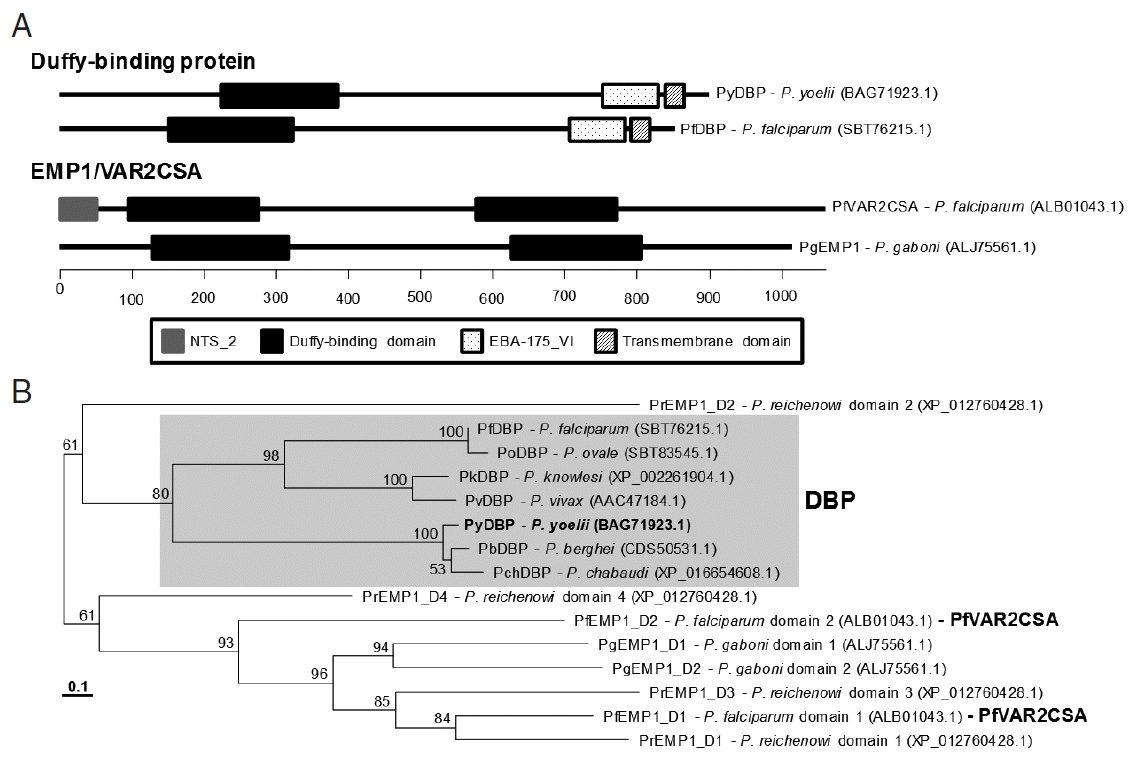
The VAR2CSA gene was screened in the genome of various P. yoelii stains by tBLASTn program using the aa sequence of P. falciparum VAR2CSA (partial, ALB01043.1) as a query. The sequence retrieved an mRNA transcript putatively encoding Duffy-binding protein (DBP; PY04764 of 17XNL strain, PY17X_1337400 of 17X strain and PYYM_1334400 of YM strain; E-value=7e-13). The 5′- and 3′-region sequences of PY04764 (2361 bp) differed from those of PY17X_1337400 and PYYM_1334400 (2559 bp; data not shown). The nucleotide sequence of homologous gene, which was empirically determined from the genome of 17XNL strain (Supplementary Fig. 1), was found to be identical to those of PY17X_1337400 and PYYM_1334400. Therefore, there might have been some fault during determination of PY04764 sequence. The genomic version of the P. yoelii DBP gene (PyDBP, 3,228 bp) was comprised of 5 exons and 4 intervening introns.
BLASTp search of the GenBank plasmodial databases using the PyDBP sequence retrieved numerous DBPs from the rodent and primate malaria databases including P. falciparum DBP (SBT76215.1) (identity>33%, E-value<2e-43). However, the orthologs of P. falciparum VAR2CSA, named erythrocyte membrane protein 1 (EMP1), were detected only in the proteome databases of P. reichenowi and P. gaboni (identity>35%, E-value<6e-23). The DBP and EMP1/VAR2CSA protein groups possessed the well conserved Duffy-binding domain, although the domain was duplicated in the EMP1/VAR2CSA (Fig. 3A). P. reichenowi EMP1 (XP_012760428.1) had 4 different Duffy-binding domains. Other group-specific domains such as EBA-175_VI and NTS-2 were further found in the proteins of each group. In a phylogenetic tree, the Duffy-binding domains comprising DBP formed a tight clade, which were clearly separated from those consisting of EMP1/VAR2CSA (Fig. 3B).
The plasmodial orthologs of LSA1 were screened with the amino acid sequence of P. reichenowi LSA1 (CD064792.1) against Plasmodium databases in PlasmoDB. However, matches with statistical significance were detected only in P. falciparum (PF3D7_1036400.1-p1 of 3D7 strain and PFIT_1035600.1-p1 of IT strain; E-value<7e-155). Therefore, LSA1 gene appeared to have evolved specifically in the genomes of P. falciparum and P. reichenowi.
Protective immune responses in P. yoelii 17XNL-challenged mice
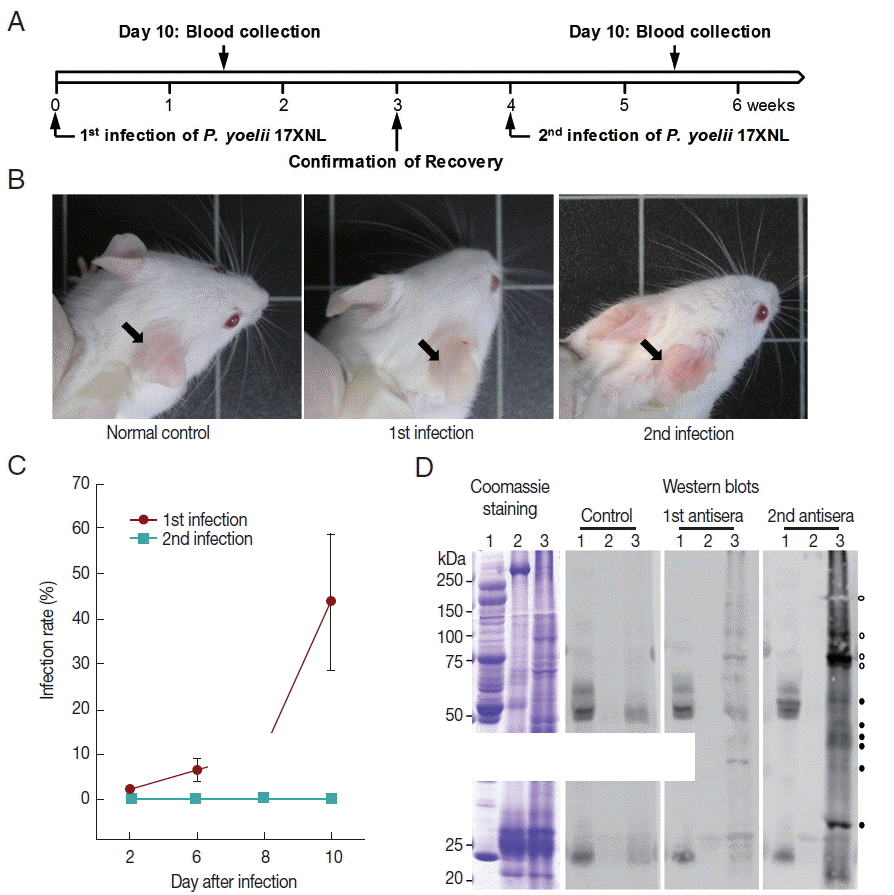
The ICR mice (n=6) were challenged with the non-lethal P. yoelii 17XNL and monitored over the course of infection for parasitemia (Fig. 4). The percent parasitemia in the peripheral blood was increased from 2.03%±0.403 at 2 days to 43.82%±15.146 at day 10 post-infection. The peak parasitemia at day 10 was gradually decreased to a background level and finally, complete convalescence from malaria was confirmed at week 3 after the primary challenge. The mice ears, which had turned to be pale during the infection with high parasitemia, recovered their original color (Fig. 4B). Interestingly, the parasite was rarely detected in the peripheral blood, when the recovered mice were secondary challenged with an identical dose of P. yoelii 17XNL (n=6; Fig. 4A–C). The mice ears also maintained the normal pinkish color (Fig. 4B). Western blot analysis of P. yoelii proteins with a pooled serum collected from the primary and secondary infected mice (n=6) demonstrated that multiple proteins invoked humoral immune response to generate specific antibodies (Fig. 4D). Some of the specific antigen-antibody reactions were uniquely detected in the secondary infected serum (closed circles), while the others were found to be intensified probably due to the repeated challenge of identical antigens in the re-infected mice (open circles).
Antigenicity of recombinant MSP1 and CSP proteins
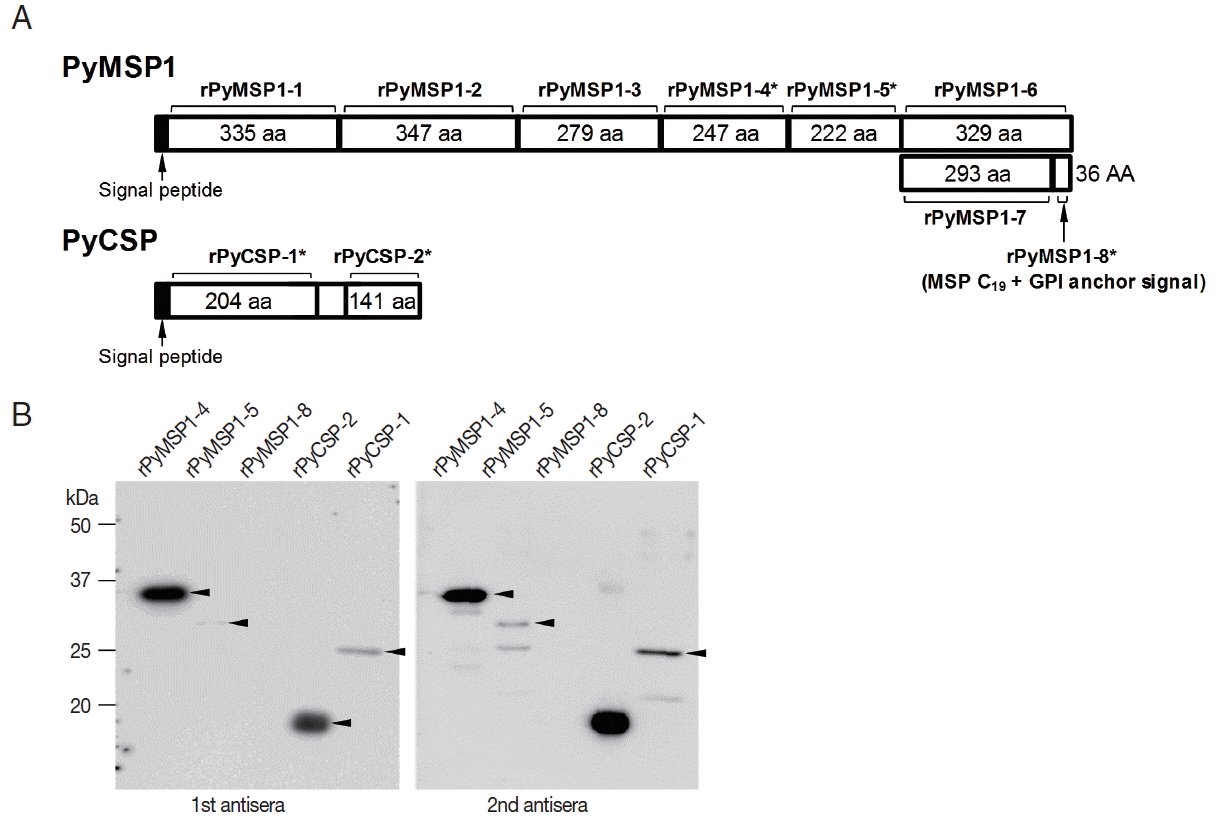
The coding DNA sequences of P. yoelii MSP1 and CSP genes were segmentally cloned to generate recombinant fragments of these proteins (Fig. 5A and Supplementary table 1). Of the 10 gene segments transformed into E. coli cells, 5 gene segments were successfully induced by IPTG (3 rPyMSP1 and 2 rPyCSP segments; recombinant fragments marked with asterisks). The recombinant proteins were purified and reacted with the P. yoelii-specific mouse antisera. As shown in Fig. 5B, the rPyMSP1-4 and rPyCSP-2 fragments exhibited strong positive signal, while rPyMSP1-5 and rPyCSP-1 reacted weakly with the mouse sera. All the signal intensity was slightly increased in reactions with sera from the secondary infection. However, no significant reaction was observable between the mouse antisera and rPyMSP1-8 containing C19 fragment, which had been known to act as a potent B-cell epitope in P. falciparum MSP1.
DISCUSSION
Plasmodial proteins that are specifically expressed during erythrocyte stage have been targeted for the development of protective vaccines [7,25,26]. Up to date, several proteins expressed on the surface of the merozoites and infected red blood cells such as merozoite surface proteins 1/2/3 (MSP1/2/3), serine-repeat antigen, erythrocyte binding antigen, glutamate-rich protein (GLURP) and apical membrane antigen 1(AMA1) have been developed as potent vaccines [7]. MSP1 engaging in the initial attachment of merozoites to erythrocytes is a leading erythrocyte-stage vaccine candidate, which has been most extensively studied in the relevant fields [27]. Both of the N-terminal and C-terminal fragments of MSP1 have been found to be effective for the generation of immune response and protection against malaria [28,29], although the C-terminal fragment appeared to exhibit larger effect than the N-terminal counterpart [26]. The protective immunity has also been examined with rodent model using the C-terminal fragment of MSP1 protein expressed in P. yoelii [16,30]. However, it was unclear whether the C-terminal fragment of native PyMSP1 invoke generation of specific antibody during P. yoelii infection, since no significant immune reaction was detected against the C-terminal C19-containg fragment of PyMSP1 (rPyMSP1-8; Fig. 5). In addition, the middle fragment of PyMSP1 A (rPyMSP1-4) also exhibited strong reactivity against mouse sera infected with the parasite.
CSP is a sporozoite stage-specific protein that expresses a unique immunodominant epitope recognized by immunized or repeatedly infected hosts [24,31,32]. The plasmodial protein contains N- and C- terminal regions of relatively low immunogenicity, which flank a central region with tandemly repeated aa units. The central repeats differ from species to species and have been targeted as vaccine candidates for protective immunity against malarial infection due to their highly immunogenic properties [23,33,34]. The central repeats of CSP encoded in the P. yoelii genome were ‘QGPGAP’ and ‘PPQQ’, which were separated by a di-amino acid sequence ‘QE’. The repeat units were well-conserved in the CSP orthologs identified in different P. yoelii strains (Fig. 2). The significant length polymorphism observed among these orthologs were mainly resulted from the shrinkage/expansion of the former ‘QGPGAP’ repeat. Meanwhile, the repeats were ‘PPPPNPND’ and ‘PQPQ’ in CSP protein of P. berghei, while the sequence of former unit was slightly deviated to ‘PPPPNAND’ and ‘PAPPNAND’ in the downstream regions. More recent investigations demonstrated that the N-terminal region of CSP proteins (double underlined sequence in Fig. 2) can also be applicable for the development of protective vaccines, even though its protective potential was relatively weak compared to that of the central repeat [24,34]. In P. yoelii system, the specific mouse antisera reacted strongly with the C-terminal fragment of CSP (rPyCSP-2), but not with the N-terminal fragment containing the ‘QGPGAP’ repeat (rPyCSP-1) (Fig. 5B). Taken together, these facts may reflect the collaborated actions between multiple regions of CSP proteins, which invoke specific immune response(s) against the parasite infection, are uniquely evolved among Plasmodium species.
The adhesion of infected erythrocytes to vascular endothelium or placenta is the key event in the pathogenesis of severe malaria induced by P. falciparum infection. By binding to receptors in the vascular bed, the parasite avoids being filtered through spleen, where they are removed from the circulation [35,36]. The plasmodial EMP1 proteins are adhesion molecules mediating the interaction between infected erythrocytes and the host endothelium. Of the multiple paralogous EMP1 proteins, VAR2CSA is a unique variant of EMP1 that is expressed in pregnant women and is responsible for the adhesive binding in the placenta. The VAR2CSA protein interacts with a low-sulfated form of chondroitin sulfate A attached to proteoglycans in the intervillous spaces of the placenta [37–40]. Protective immunity against the pregnancy-related placental malaria is acquired over multiple pregnancies and thus, VAR2CSA can be a major target for the development of vaccine against placental malaria [39,41]. In this study, VAR2CSA homologs are detected only in P. reichenowi and P. gaboni, but not in other primate and rodent malaria (Fig. 3). Currently, it is uncertain whether the EMP1 protein plays the adhesive role, similar to the VAR2CSA protein of P. falciparum, in rodent malaria. Nevertheless, the functional analog(s) of VAR2CSA is likely to be expressed in rodent malaria, considering many features that are commonly observed in the murine and human placental malaria [42]. Screening of the functional analog(s) in murine system is necessary to use the rodent malaria as an effective experimental model of human placental malaria.
To date, numerous recombinant proteins and crude blood-stage antigens of plasmodial species have been applied in the experimental investigations of protective immunity against malaria using model animals [14,26,43]. The target proteins mixed with various adjuvants were injected into experimental animals in a large amount to invoke potent immune response in these studies [14]. However, the ability to stimulate host immune system for the generation of specific protective antibodies appears to be quite different among vaccine candidates in practical patients [26]. Therefore, there have been more recent efforts to overcome/compensate the clinical limitation of previously established vaccine candidates by selecting immunogenic antigens against naturally acquired antibodies [43–45]. Several protein bands expressed in the blood-stage P. yoelii 17XNL were highly reactive with specific antibodies in experimental mice sera recovered from the primary challenge of the parasite. The low or negligible parasitemia observed in these mice might result from the neutralizing action of the protective antibodies (Fig. 4). In addition to the generation of recombinant proteins characterized in this study for protective test, experiments for identification and cloning of the immunogenic antigens shown in Fig. 4D via mass spectrometry and following Mascot analyses are under preparations. Vaccination effects of the 17XNL proteins will also be further examined in mice challenged with the lethal P. yoelii 17X strain. The results obtained by the near-future studies might provide the necessary information and means to characterize additional vaccine candidate antigens, selected solely on their ability to produce the protective antibodies.