Echinococcus granulosus sensu lato (s.l.) is a causative agent of cystic echinococcosis or cystic hydatid disease in humans and domestic and wild animals [1]. This disease is a serious health problem in countries associated with poverty and poor hygiene practices, particularly livestock raising [2]. Until recently, the taxonomy of E. granulosus s.l. spp. has been controversial because of the limited availability of morphological diagnostic characters in both adult and larval stages [3]. The incomplete development of metacestodes in aberrant hosts has further limited the identification of causative organisms. In the 1990s, an Australian research group [4–6] conducted molecular taxonomic analyses of E. granulosus s.l. using short mitochondrial DNA sequences of genes for cytochrome c oxidase subunit 1 (cox1) and NADH dehydrogenase subunit 1 (nad1). The analyses showed that Echinococcus multilocularis, Echinococcus vogeli, and Echinococcus oligarthra are distinct from each other and that E. granulosus s.l. can be divided into genotypes G1–G8 [7]. These genotypes correspond to the following strains: E. granulosus sensu stricto (s.s.) G1 (sheep strain), G2 (Tasmanian sheep strain), G3 (buffalo strain); Echinococcus equinus G4 (horse strain); Echinococcus ortleppi G5 (cattle strain); Echinococcus canadensis G6 (camel strain), G7 (pig strain), and G8 (cervid strain) [3,8]. Currently, these genotypes are used as standards for molecular classification when considering the epidemiology of cystic echinococcosis [9]. Another E. canadensis G9 (unidentified strain) and G10 (Fennoscandian cervid strain) were subsequently characterized [10,11], while the status of G9 remains unclear [3,8].
Although entire or partial gene sequencing is effective for determining genotypes, specialized equipment is required to conduct these costly and time-consuming procedures. In many developing countries, the supply of such equipment is insufficient, and thus PCR-RFLP is frequently used [12,13]. In fact, most countries in which this parasite is prevalent are economically underdeveloped, and thus a simple PCR-RFLP method is necessary. However, PCR-RFLP results must be compared with previously reported PCR-RFLP results. Using this method, some under-digested DNA fragments may be confused with fully digested fragments, limiting the interpretation of PCR-RFLP results. To overcome this limitation, we developed a PCR-RFLP algorism involving only 2 or 3 restriction enzymes that produce only 1 or 2 fragments in PCR-RFLP, which is highly helpful for improving the quality of the results.
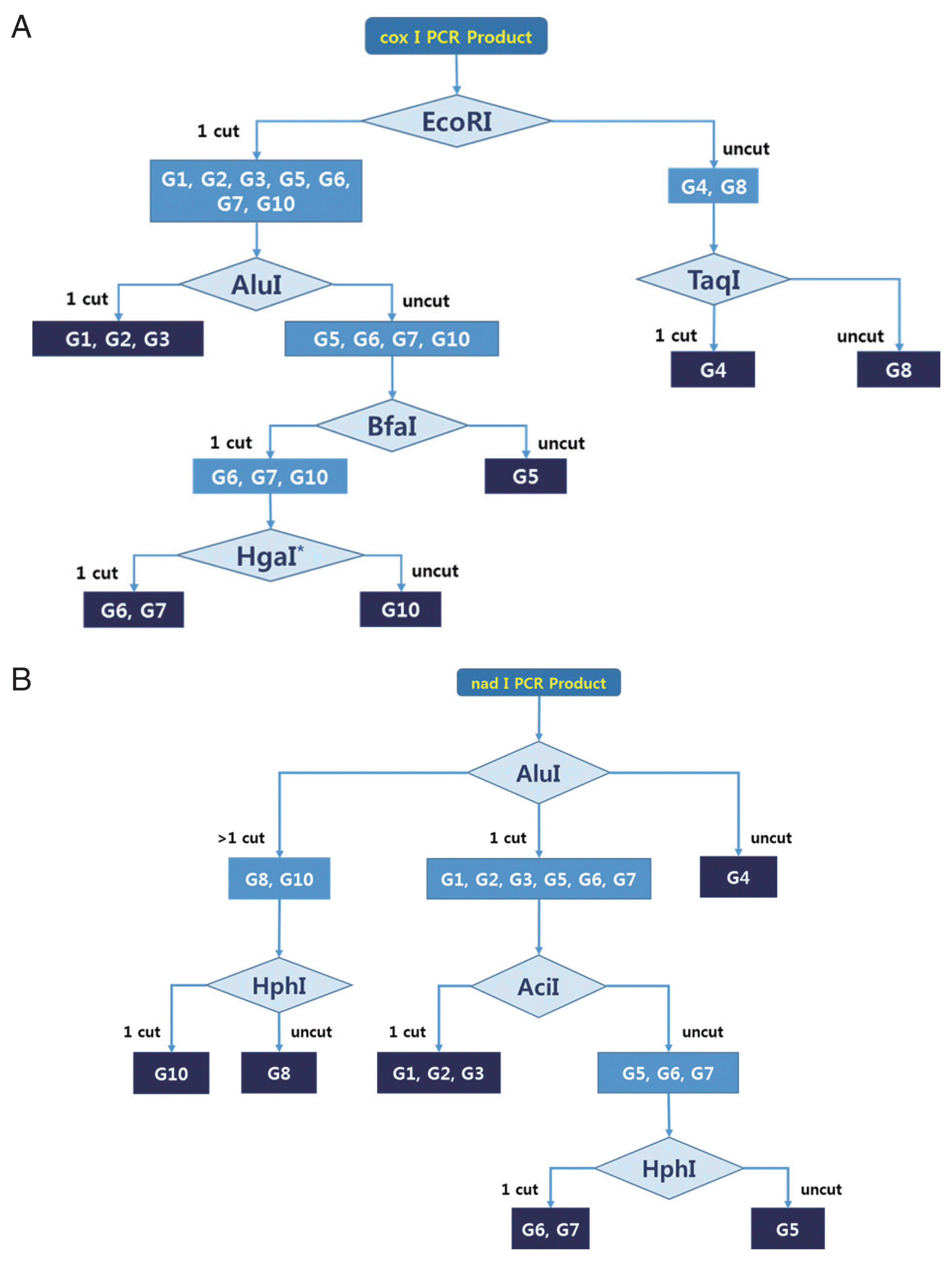
The cox1 and nad1 sequences from 9 E. granulosus s.l. genotypes (G1–G10, except G9) were downloaded from GenBank (https://www.ncbi.nlm.nih.gov/nucgss). After multiple alignment (MEGA software version 6), we searched for new PCR target lesions that could produce 1 or 2 fragments after restriction endonuclease digestion using NEBcutter version 2.0 (http://nc2.neb.com/NEBcutter2/). The PCR primer sets for cox1 and nad1 are constructed like as cox-f [CC(C/T) GG(A/G) TTT GGT(A/G) T(A/T) ATT], cox-r [ATC(A/G) TG(C/T) AA AA(C/T) ATT ATC], nad-f [A(G/A)(A/T) TTC GTA AGG G(G/C) C CTA ATA], and nad-r [(A/T)CC(A/T) CT AAC(T/C) AA TTC ACT TTC]. We predicted hypothetical PCR-RFLP fragments based on various endonuclease recognition sites, and selected appropriate restriction enzyme sets for each cox1 and nad1 PCR RFLP algorism. We used more than 10 restriction enzymes for hypothetical RFLP, as it was not possible to identify all genotypes using only 1 enzyme. Our goal was to develop an algorism that could identify all genotypes using the fewest enzymes. Most genotypes could be distinguished by using a specific restriction enzyme (Supplementary Fig. 1). However, we could not distinguish between the G1–G2, G1–G3, G2–G3, and G6–G7 genotypes using cox1 and nad1 PCR-RFLP in this study. Finally, we determined the best algorism for 2 mitochondrial genes (cox1, nad1) by comparing band patterns (Fig. 1). To evaluate the algorism, unidentified genomic DNA was isolated from 3 Echinococcus protoscoleces. Three hydatid cyst fluids were collected from patients who underwent medical treatment at Samarqand Hospital in Uzbekistan. The protoscoleces were obtained after centrifugation for 5 min at 2,000 ×g. Total genomic DNA was extracted using the QIAamp DNA Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer instructions. The genomic DNA was eluted in 200 μl of water and stored at −20°C until PCR amplification. The extracted genomic DNA was amplified using noble primer sets for 2 mitochondrial DNA genes. PCR was carried out in a final volume of 50 μl containing 5 μl genomic DNA, 10X EF-Taq Reaction Buffer (25 mM MgCl2 mixed), 2 μl of 10 mM dNTPs, 5 μl of 10 pmol of each primer, and 1 μl (2.5 U) of DiaStarTM EF-Taq DNA Polymerase in a thermocycler (Takara, Shiga, Japan).
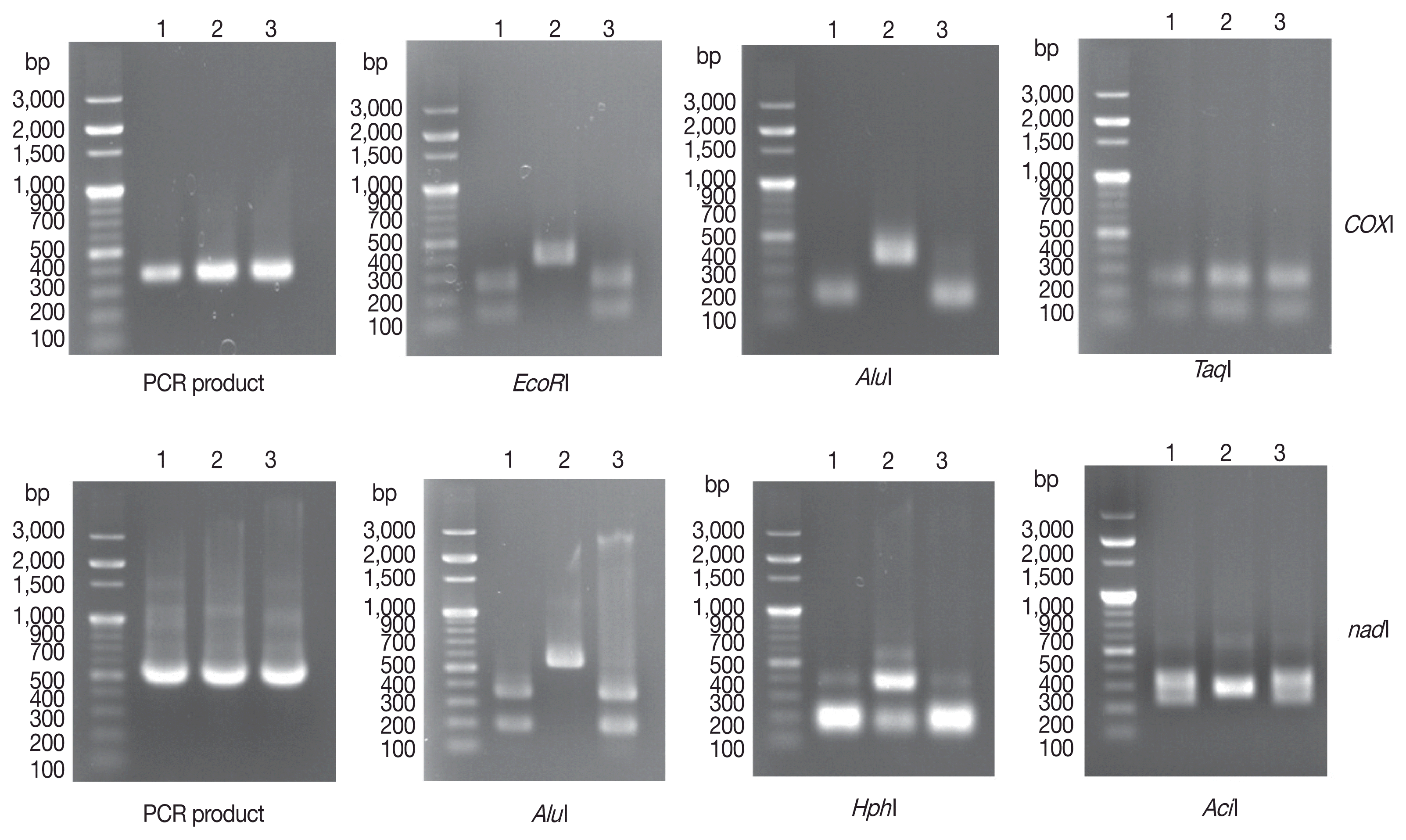
The following temperature profile was used for cox1 amplification: initial denaturation at 94°C for 30 sec; followed by 35 cycles of denaturation at 94°C for 30 sec, annealing at 42°C for 45 sec, and elongation at 72°C for 35 sec. This was followed by final extension at 72°C for 10 min. The following temperature profile was used for nad1 amplification: initial denaturation at 94°C for 30 sec; followed by 35 cycles of denaturation at 94°C for 30 sec, annealing at 45°C for 45 sec, and elongation at 72°C for 35 sec. Final extension was performed at 72°C for 10 min. PCR products were confirmed on 1.0% agarose gels under ultraviolet (UV) light. The PCR products were digested for 2–3 hr at 37°C by 4 restriction enzymes (AluI, EcoRI, AciI, and HphI) and at 65°C by the restriction enzyme TaqI. Restriction fragments were separated on 1.5% agarose gels and photographed under UV light. After 1% agarose gel electrophoresis of cox1 and nad1 PCR products, we obtained 350-base pair (bp) (expected size 366 bp) cox1 PCR products and 500-bp (expected size 530 bp) nad1 PCR products (Fig. 2) from 3 human Echinococcus sample. After digestion of cox1 gene PCR products with EcoRI, 2 PCR-RFLP patterns were observed. The genotypes of the 2 samples (unknowns 1 and 3) were likely G1, G2, G3, G5, G6, G7, or G10, while that of unknown 2 was likely G4 or G8. After AluI digestion, we found that unknowns 1 and 3 were included in E. granulosus s.s (G1, G2, and G3 genotypes). Finally, we found that unknown 2 was included in E. equinus (G4 genotype) based on TaqI digestion. For the nad1 PCR-RFLP genotyping algorism, the same results were observed for 3 different restriction enzymes (AluI, HphI, and AciI) (Fig. 2).
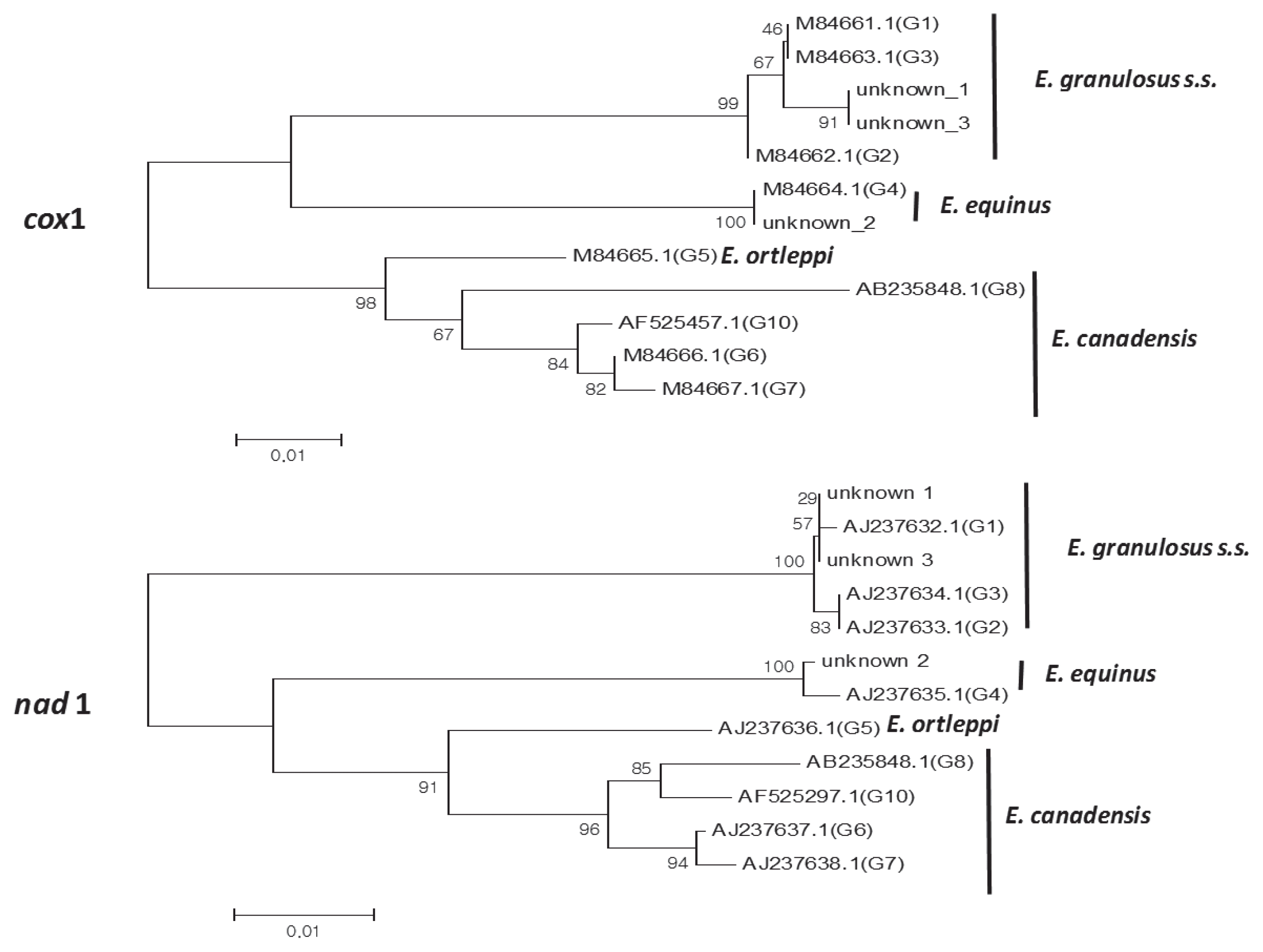
To confirm the PCR-RFLP algorism, nad1 and cox1 PCR products of unidentified genotypes were sequenced after purifying the PCR products with ExpinTM PCR SV (GeneAll Biotech, Seoul, Korea). Sequences and the reference E. granulosus s.l. genotype sequences were subjected to multiple alignment and phylogenetic distances were calculated using BLAST (http://www.ncbi.nlm.nih.gov/) and MEGA software version 6. Phylogenetic trees were constructed using the neighbor-joining method in MEGA software version 6.0. DNA sequencing yielded consensus sequences of the cox1 and nad1 genes. Key reference sequences for genotyping were used according to previous studies [14,15]. Sequences analysis of cox1 PCR products using alignment and phylogenetic analysis with reference sequences indicated that unknowns 1 and 3 were very similar to the sequences of G1 (99% identity), G2 (99% identity) and G3 (99% identity). The entire sequence of cox1 PCR product of unknown 2 was identical to that of the G4 genotype (Fig. 3). For nad1 gene sequence analysis, although there were some insignificant differences, we obtained similar results as those obtained for cox1 (Fig. 3).
Based on morphological characteristics, 4 species are currently recognized in the genus Echinococcus: E. multilocularis, E. vogeli, E. oligarthra, and E. granulosus s.l. Other species such as E. shiquicus, E. felidis, E. ortleppi, E. equinus, E. canadensis and E. intermedius are currently classified as E. granulosus s.l. [3,8,16,17]. We constructed practical algorism for rapid genotyping of E. granulosus s.l. isolated from human and domestic animals using only 1–3 restriction enzymes. This method is less cost and more rapid than sequencing analysis. Although G1, G2, and G3 were not distinguished by this algorism, these genotypes have been recognized as the same species, E. granulosus s.s. [15,18,19]. The algorism distinguished G8 and G10 from the G6/G7 complex, but E. canadensis G6 and G7 could not be distinguished. Previous studies were also unable to separate the G6/G7 complex using mitochondria DNA, ITS1, and 18S ribosomal DNA sequencing [8,20].
There were some limitations to this study. The algorisms were developed for only 9 genotypes (G1–G10, except G9) of E. granulosus s.l. and not for E. felidis, E. multilocularis, E. shiquicus, E. oligarthra, and E. vogeli, which use wild animals as their intermediated hosts [8]. However, among these species, only E. multilocularis is known to commonly infect humans and is not found in domestic animals. Additionally, the morphological characteristics of their hydatid cysts are quite different from those of E. granulosus s.l. [20]. In these cases, genetic classification may be unnecessary for specific identification. Interestingly, we identified the first human infection case of E. equinus (Fig. 3). No human infections by the G4 genotype have been detected previously [21].
Supplementary materials
Supplementary Fig. 1
Schematic representation of hypothetical RFLP of E. granulosus s.l. species using four restriction enzymes. (A) The hypothetical pattern of bands using PCR product of coxI by AluI, EcoRI, TaqI, BfaI, and HgaI. (B) The hypothetical pattern of bands using PCR products of and 1 gene by AluI, HphI, and AciI. The 100 bp DNA Ladder Marker (Enzynomics, Daejeon, Korea) was used as a size marker.