INTRODUCTION
Toxoplasma gondii is an important obligate intracellular parasitic protist that can virtually infect all the warm-blooded animals including humans, causing serious clinical diseases and noticeable veterinary problems especially for pregnant women and immunocompromised patients [1,2]. In addition, so far there are no ideal drugs for the therapy of toxoplasmosis [3]. Therefore, an effective and safe vaccine is urgently needed to prevent and control the infection of this parasite both in medical and veterinary circumstances.
Heat shock proteins (HSPs) with wide distribution are the highly conserved molecular chaperones, which can keep the proteins from mutation, misfolding, inaccurate modification, and the acute influence of environment or chronic insult etc; HSPs are consecutively expressed during the growth process of cell cycles and play an important regulatory role in the protein folding/refolding, repair, degradation and the intracellular transportation [4–6]. In addition to being consecutively induced, HSPs can be also triggered by a series of physiological, pathological or environmental factors and are relevant to various clinical diseases such as stress, infection, autoimmunity and cancer [7–9]. HSPs may have potentially experimental and/or clinical applications as the molecular biomarkers for etiological diagnosis, therapeutic targets for disease treatment, or vaccine candidates for epidemic prevention in animals and human beings.
As an important member of the HSP family, T. gondii HSP60 was located in mitochondria and acts as a molecular chaperone that participates in stage-specific induction in the respiratory pathway of this parasite, mediating the activation of antigen presenting cells by stimulating inflammatory factors and inducing initial immune responses, which suggests that HSP60 may be a good DNA vaccine candidate for the prevention and control of T. gondii [10,11]. In this study, pVAX-HSP60 DNA vaccine was constructed using pVAX I vector and its protective effects against the acute and chronic infections of T. gondii was evaluated in Kunming mouse.
MATERIALS AND METHODS
Animals and ethics statement
Six-week-old female Kunming mice of specific-pathogenfree (SPF) grade were used in the present study. Kunming mice have clear genetic backgrounds, immunological and hematological indexes, and are the most commonly used laboratory animals for biological and biochemical studies in China [12,13]. A number of previous studies have shown that Kunming mice are quite susceptible to T. gondii infection, and they serve as ideal model for vaccination studies against T. gondii infection [13]. Kunming mice were purchased from the Laboratory Animal Center, Lanzhou Institute of Biological Products (Lanzhou, China), and all the animal procedures in the study were approved by the Animal Ethics and Administration Committee of Lanzhou Veterinary Research Institute, Chinese Academy of Agricultural Sciences (Approval No. LVRIAEC2012-011).
Parasites
Two strains of T. gondii (RH and PRU) saved in the Parasitology Department of Lanzhou Veterinary Research Institute were used in the present study. RH tachyzoites (Type I) maintained by serial passage in African green monkey kidney (Vero) cell monolayers were collected, washed and re-suspended as described previously [13]. Cys ts of PRU strain (Type II) sustained through monthly passage were separated from brains of the orally infected female Kunming mice. The purified RH and PRU parasites were quantified for preparation of Toxoplasma lysate antigen (TLA) and challenge of immunized Kunming mice [13].
Construction of recombinant plasmid
Total RNA of T. gondii RH tachyzoites was extracted along the instruction of E.Z.N.A.® Total RNA Kit I (Omega Bio-Tek, Norcross, Georgia, USA) and used to construct pVAX-HSP60 DNA vaccine. A pair of specific primers (forward primer: 5′-GGGGTACCATGCTTGCCCGCGCTTCAGC-3′; reverse primer: 5′-AAGGAAAAAAGCGGCCGCCTAGTACATGCCTCCCATGCCGC-3′) were designed to duplicate the full-length coding sequence of Toxoplasma HSP60 gene (GenBank XM_002367081), in which 2 restriction enzyme sites (KpnI and NotI) were underlined.
An RT-PCR amplification was performed as the guidance of PrimeScript® One Step RT-PCR Kit (Takara, Dalian, China): reverse transcription at 50.0°C for 30 min; initial denaturation at 94.0°C for 2 min followed by 35 cycles composing of 94.0°C for 1 min, 60.4°C for 45 sec and 72.0°C for 2 min; the final extension step at 72.0°C for 10 min. The products were purified and subsequently inserted into pMD18-T vector (Takara), forming pMD-HSP60 recombinant vector. The HSP60 sticky fragments were cut from pMD-HSP60 vector using KpnI and NotI, purified with gel extraction and subsequently inserted into the corresponding site of pVAX I vector through T4 DNA ligase (Takara), generating the recombinant plasmid pVAX-HSP60. The concentration of pVAX-HSP60 was determined using spectrophotometer at OD260 and OD280. Sequence accuracy was further confirmed by PCR, DNA sequencing and double-enzyme digestion.
Immunofluorescence
Human embryonic kidney 293 cells (HEK293) were used to confirm that whether the HSP60 recombinant protein was expressed in eukaryotic cell as previous description [14]. In brief, HEK293 cells grown in 6-well plate were transfected with 10.0 μg pVAX-HSP60 endotoxin-free plasmid using lipofectamine 2000 (Invitrogen, Carlsbad, California, USA) as instructed by the manufacturer. Three days post-transfection, the samples were fixed, permeabilized, blocked and incubated with goat anti-Toxoplasma serum at 37.0°C and subsequently secondary antibody at room temperature (RT) with darkness for 60 min. The dilutions of primary and secondary antibodies were as follows: the serum stored in our laboratory, 1:50; Alexa Fluor® 488-AffiniPure Donkey Anti-Goat IgG (H+L) (Jackson ImmunoResearch Laboratories Inc, West Grove, Pennsylvania, USA), 1:1,000. Group transfected with pVAX I vector was used as the control. The fluorescent images were recorded by fluorescence microscope (Olymplus IX 51, Olympus, Tokyo, Japan) with camera D7000 (Nikon, Tokyo, Japan).
Immunization and challenge
The female Kunming mice were randomly separated into 4 groups (42 mice per group, shown in Table 1), and their immunization and challenge were performed as the previous approaches with some modifications [15]. Three groups of Kunming mice were respectively immunized with 100 μg pVAX-HSP60, 100 μg pVAX I or 100 μl of sterile PBS for 3 times with 2-week interval (shown in Table 1). The rest group without any treatment was used as blank control. Three mice each group were handled by ophthalmectomy for accessing humoral immunity, among which spleens of the 3 mice before challenge were also aseptically removed to estimate the cell-mediated immunity such as lymphocyte proliferation, cytokines and flow cytometric analyses.
To further determine whether the immune responses induced by pVAX-HSP60 protect Kunming mice against the acute and chronic Toxoplasma infections, 15 mice in all the groups were intraperitoneally inoculated with 1,000 tachyzoites of RH strain, and their survival time was daily recorded until all the mice were dead. Another 15 mice were challenged with 10 cysts of PRU isolate by oral, and the total number of PRU tissue cysts in their whole brains were counted as the previous description at 30 days post-challenge for evaluating the immune response against the chronic infection (shown in Table 1) [13]. All the experimental and blank control samples were analyzed in triplicate.
Antibody detection
SBA ClonotypingTM System/HRP Kit (Southern Biotech, Birmingham, Alabama, USA) and ELISA plates pre-coated with soluble rHSP60 protein (preserved in our laboratory) were used to determine the levels of specific anti-Toxoplasma antibodies (total IgG, IgG1, and IgG2a) in the serum samples. Reference to the kit instruction, ELISA plates were coated with 1.0 μg rHSP60 protein per well overnight at 4.0°C, washed with 300 μl of PBS containing 0.05% Tween 20 (PBST) per well for 3 times and subsequently blocked with 100 μl of 1.0% BSA/PBS for 1 hr. 100 μl of serum sample was then added to the empty well and incubated at RT for 1 hr with gentle shaking. After washing with PBST, the well was incubated at RT for 1 hr with 100 μl of HRP-conjugated anti-mouse Ig, IgG1 or IgG2a diluted in 1:500, respectively. The color development was performed with 100 μl of substrate solution containing 1.05% citric acid, 0.03% H2O2 and 0.03% ABTS provided by the kit. The optical density was measured by ELISA reader (Bio-Tek ELX800, Winooski, Vermont, USA) at 405 nm after substrate addition according to the manufacturer’s instruction.
Flow cytometric analysis
Spleens of the 3 mice each group were also aseptically removed before challenge, and the single-cell suspensions were prepared using the aseptic wire mesh and erythrocyte lysis solution (Macgene, Beijing, China) [13]. After counting using 0.04% Trypan Blue dye, 1×105 cells in the suspension were diluted in 100 μl of 2% FBS/RPMI-1640 medium and incubated at 4°C for 30 min in the dark with 0.5 μl of PE anti-mouse CD3e, 0.3 μl of APC anti-mouse CD4 and/or 0.2 μl of FITC anti-mouse CD8a (eBioscience, San Diego, California, USA) referring to the manufacturer’s guidance. All the samples were washed with 2 ml of PBS, re-suspended with 300 μl of FACScan buffer (1% BSA, 0.1% NaN3) and fixed with 250 μl of 2% paraformaldehyde. The data were gathered using FACScan flow cytometer (BD Bio-sciences, San Jose, California, USA) and SYSTEM II software (Beckman-Coulter, Miami, Florida, USA). All the samples were carried out in 3 independent experiments. Cytokine detection and lymphocyte proliferation 3×105 cells of the single-cell suspension diluted in 200 μl of FBS/RPMI-1640 medium were incubated with 5 μg/ml concanavalin A (ConA, Sigma) at 37°C in a 5% CO2 incubator. The cell-free culture supernatants were collected by centrifugation for 5 min at 3,000 rpm, and their levels were quantified using pre-coated 96-well plates after 1 day for IL-2 and IL-4, 3 days for IL-10, and 4 days for IL-12p70 and IFN-γ according to the manufacturer’s instructions (Biolegend, San Diego, California, USA). The concentrations of cytokines were calculated referring to the standard curves that were constructed with the known amounts of mouse recombinant IL-2, IL-4, IL-10, IL-12p70, and IFN-γ provided by the kits. The data were harvested from 3 independent experiments.
The same number of single-cell suspension were used for lymphocyte proliferation assay in this study. Briefly, 100 μl of lymphocyte solution was spread into 96-well costar plate and subsequently incubated with isometric TLA (10 μg/ml), ConA (5 μg/ml) or medium at 37°C in a 5% CO2 incubator. Evaluation of lymphocyte proliferation was performed using stimulation index (SI) determined by 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt (MTS, 5 mg/ml, Promega) [16]. SI was defined as the ratio of the average absorption value at 570 nm of wells containing TLA-induced cells (AVTLA) to ConA-induced cells (AVConA) compared to the medium (AVM), respectively. That is, SI=(AVTLA/AVM)/(AVConA/AVM). All the samples were measured in triplicate.
Statistical analysis
All the measured data were analyzed by SPSS13.0 Data Editor (SPSS Inc., Chicago, IL, USA). Difference between the groups was calculated using student′s t-test, if their variances analyzed by F-test were equal. A result was considered as different, when P <0.05 or significantly different when P <0.01.
RESULTS
HSP60 expression in HEK293 cell
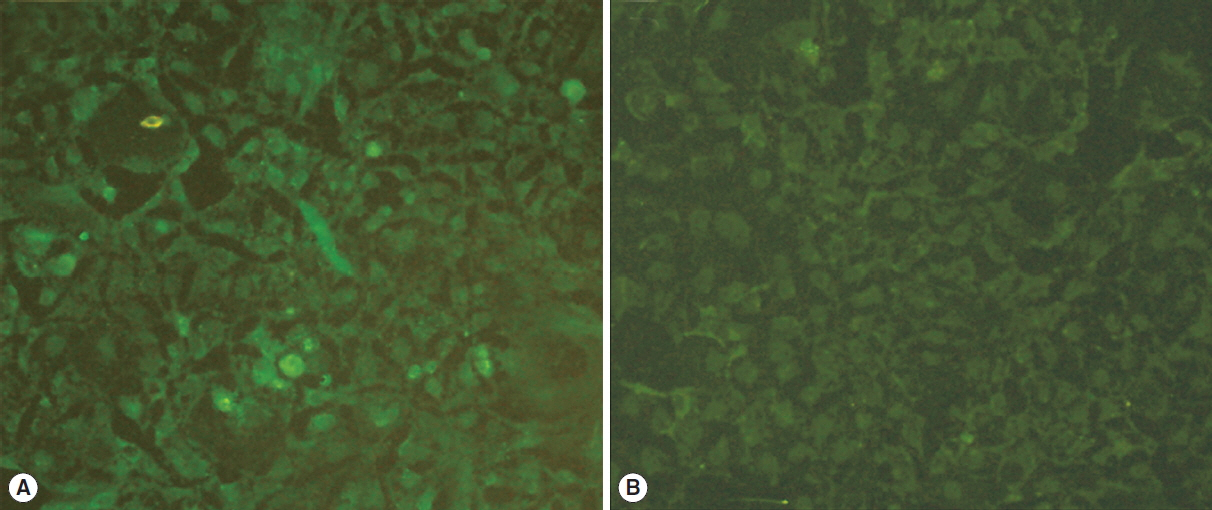
Immunofluorescence assay with goat anti-Toxoplasma antibody was performed to confirm whether pVAX-HSP60 plasmid could be expressed in HEK293 cell in this study, the results of which showed that the specific green fluorescent substances were detected in the cells transfected with pVAX-HSP60, but no fluorescence in the control group (shown in Fig. 1), suggesting that Toxoplasma HSP60 recombinant protein can be expressed in eukaryotic cells.
Antibody responses in the immunized mice
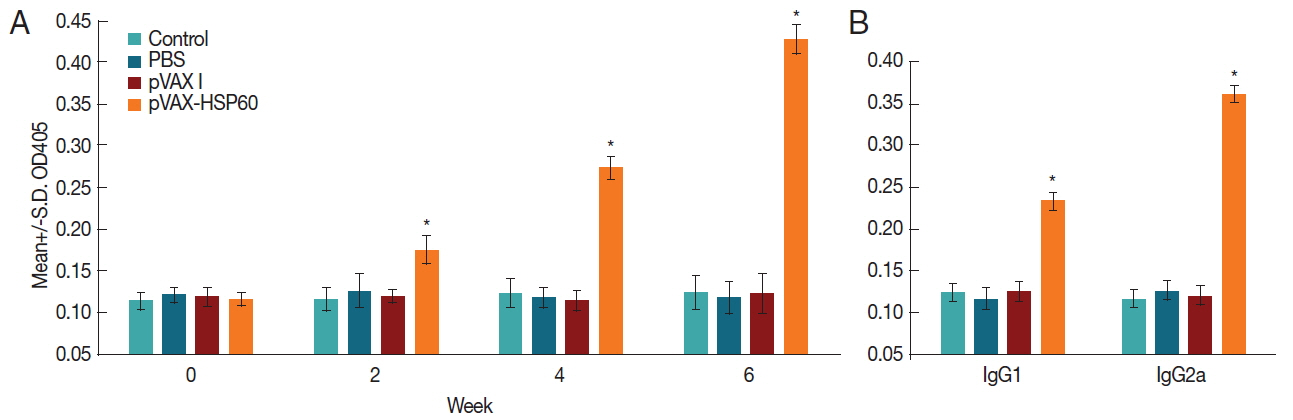
The specific anti-Toxoplasma antibodies (total IgG, IgG1, and IgG2a) in the serum samples were examined through coating with soluble rHSP60 protein and subsequent detection using SBA ClonotypingTM System/HRP Kit to evaluate the levels of humoral immune responses induced by pVAX-HSP60 immunization. The data showed that the levels of antibodies including total IgG, IgG1, and IgG2a were all significantly elevated in the experimental group in comparison with the control groups (pVAX I, PBS, and blank control) (Fig. 2), and the titers of total IgG in the samples increased with serial pVAX-HSP60 immunizations, suggesting that the humoral and Th1 type immune responses were both elicited.
Cellular immune responses
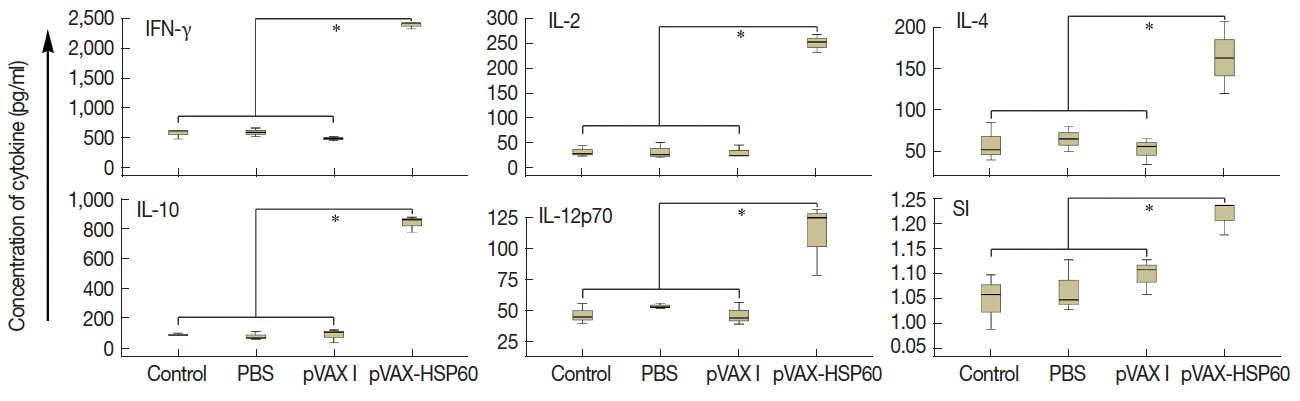
Spleen cells in Kunming mice were incubated with ConA, and 5 kinds of cytokines (IL-2, IL-4, IL-10, IL-12p70, and IFN-γ) were measured in the cell-free culture supernatants. As shown in Fig. 3A, levels of cytokines in group pVAX-HSP60 were all significantly increased in contrast to that of controls (P <0.05).
The proliferation of splenocytes that were separated at 2 weeks after the third immunization was also determined using MTS. The results showed that SI of the mice immunized with pVAX-HSP60 (1.22±0.03) was significantly higher than that of pVAX I (1.10±0.04), PBS (1.07±0.05) and blank control (1.05±0.06) (P <0.05) (Fig. 3B).
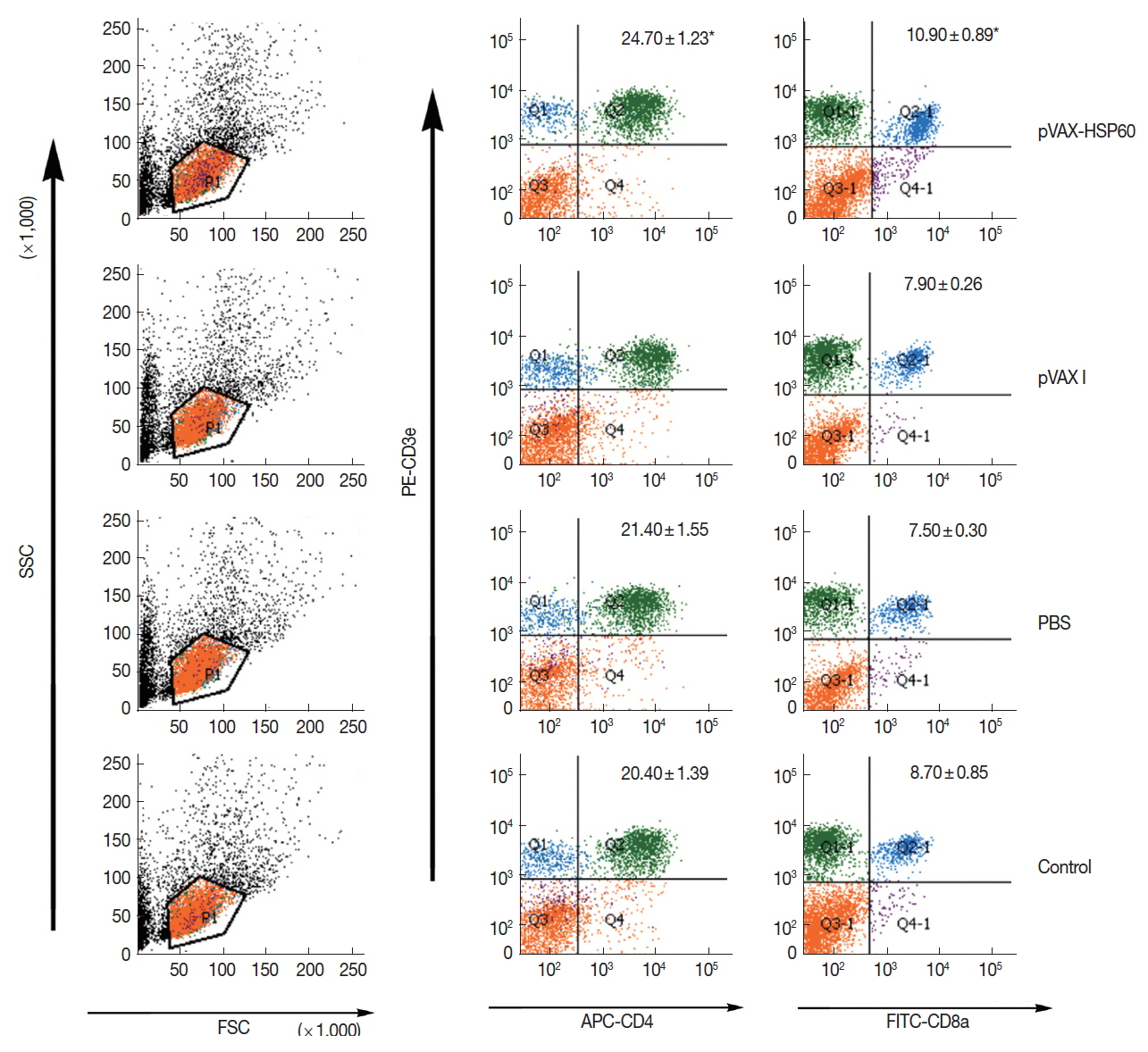
The flow cytometry analyses revealed that the percentages of CD3e+CD4+ and CD3e+CD8a+ T cells performed as means +/− S.D. were marked (Fig. 4). It was found that CD3e+CD4+ and CD3e+CD8a+ T cells in pVAX-HSP60-immunized group were significantly increased in comparison to that of controls (P <0.05). It was indicated that Th1- and Th2-type cellular immune responses were both triggered through the successive DNA immunizations.
Immune protection against T. gondii infections
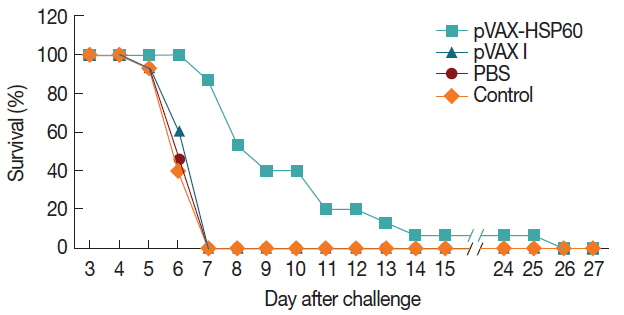
To assess whether pVAX-HSP60 immunization could produce immune protections against the acute and chronic Toxoplasma infections, all the groups of Kunming mice were challenged with 1,000 tachyzoites of RH strain or 10 cysts of PRU isolate. The survival times of mice infected with RH parasites were daily recorded until all of them were dead, and significant difference was determined in the mice immunized with pVAX-HSP60 (10.53±4.78 day) compared to that in groups of pVAX I (6.53±0.64 day), PBS (6.40±0.63 day) and blank control (6.33±0.62 day) (P <0.05) (Fig. 5).
Total number of PRU cysts in whole brain was detected at 30 days post-challenge to evaluate the protective efficacy against the chronic infection of this parasite, the results of which demonstrated that the brain cysts in Kunming mice receiving pVAX-HSP60 immunization were significantly reduced in contrast to that of controls (P <0.01; Table 2). Meanwhile, lower but not statistically different cyst loads were detected in other 2 groups (Table 2).
DISCUSSION
The intracellular protozoan T. gondii with world-wide distribution has deservedly received the basic medical and scientific attentions as it is quite capable of infecting all warm-blooded animals from avian to mammals and leads to serious congenital diseases, abortion in pregnant women and domestic animals, and Toxoplasma encephalitis in immunocompromised individuals etc. [17–19]. In consideration of no ideal medicines for the clinical therapy so far, a safe and effective countermeasure should be forwardly executed to restrict the spreading and development of this parasite [3].
In the last several years, many attempts including subunit and live-attenuated Toxoplasma strain vaccines have been made, but not feasible because of their unsatisfactory effects, expensiveness and short shelf life [20,21]. Thus the exploration of DNA vaccine has been focused under the current efforts [22,23]. In addition, the previous reports have demonstrated that DNA-embarked vaccines possessed the articulated skeleton of pVAX I vector are able to express various genes in eukaryotic cells such as Marc-145 and HEK293, and trigger strong humoral and cellular immune responses [24,25]. Therefore, the effective eukaryotic system based on pVAX I skeleton was used in this study to express the HSP60 protein of T. gondii, and its fairly strong transcription in eukaryotic cells was further demonstrated using immunofluorescence assay.
As described previously, the innate and adaptive immune responses in hosts can be both induced for resistance to toxoplasmosis during the natural invasion and artificial injection of T. gondii [26]. In addition, this parasite is capable of activating a strong Th1-type cellular immune response to protect Toxoplasma-infected host against acute death, characterized by the release of early IL-12 and IFN-γ [27]. Hence, 5 kinds of cytokines besides IL-12p70 and IFN-γ in the spleen cell-free culture supernatants, which are compactly connected with the innate immune response, were measured in the present study through stimulation with ConA [28]. The results showed that all the examined cytokines (IFN-γ, IL-2, IL-4, IL-10, and IL-12p70) in the pVAX-HSP60-immunized mice increased highly in contrast to that of the 3 controls, indicating that the innate immune response was significantly awakened via serial DNA immunizations.
As the important index of adaptive immune response during Toxoplasma infection, the titers of specific antibodies in serum were used to evaluate the development of toxoplasmosis for a long time [29]. Based on this consideration, total IgG, IgG1 and IgG2a in the mice were detected by coated with the soluble rHSP60 protein that was preserved in our laboratory. Our data showed that levels of specific antibodies in sera were all significantly elevated in the mice receiving DNA immunizations compared to the 3 controls, indicating that the successive pVAX-HSP60 stimuli triggered strong humoral immune responses in Kunming mice.
Moreover, several parameters of cellular immune responses were detected to further explore the elements beneficial for host against Toxoplasma acute and chronic infections in the study, and the proliferation of T lymphocytes separated from spleen tissues in the mice receiving pVAX-HSP60 immunization was determined using flow cytometry and MTS analyses, with the significant increase of CD3e+CD4+ and CD3e+CD8a+ T cells and the significantly elevated stimulation index in comparison with the controls, matching with the previous reports [13,30], which indicated that the cellular immune responses of Kunming mice were also evoked by immunization with pVAX-HSP60.
To assess whether the innate and adaptive immune responses stimulated by pVAX-HSP60 were beneficial for the resistance to toxoplasmosis, the residual Kunming mice were finally challenged with 1,000 tachyzoites of RH strain through intraperitoneal injection or 10 cysts of PRU isolate orally, and their survival times or reduction of brain cysts were examined in the end of all the experiments. We found that the immunizations with pVAX-HSP60 were able to significantly prolong the survival times and reduce the number of brain cysts in contrast to that in control groups. As the deficiencies of the single-gene vaccines do exist such as the incomplete protection, the exploitation of ideal vaccines against T. gondii infection will still be going on.
In conclusion, high humoral and cellular immune responses were induced byHSP60 DNA immunization, which were a main reason resulting in protection against T. gondii infections. It was suggested that HSP60 DNA can be a vaccine candidate to control and prevent spreading of T. gondii infections.