INTRODUCTION
Ticks are global obligate hematophagous ectoparasites of a wide variety of terrestrial vertebrates and significant vectors of diseases in humans, domestic animals, and wildlife [1]. Tick-borne pathogens include various microorganisms, such as viruses, bacteria, and protozoans [2,3]. Approximately 119 tick species, including 100 species of Ixodidae and 19 species of Argasidae, have been identified in China [4].
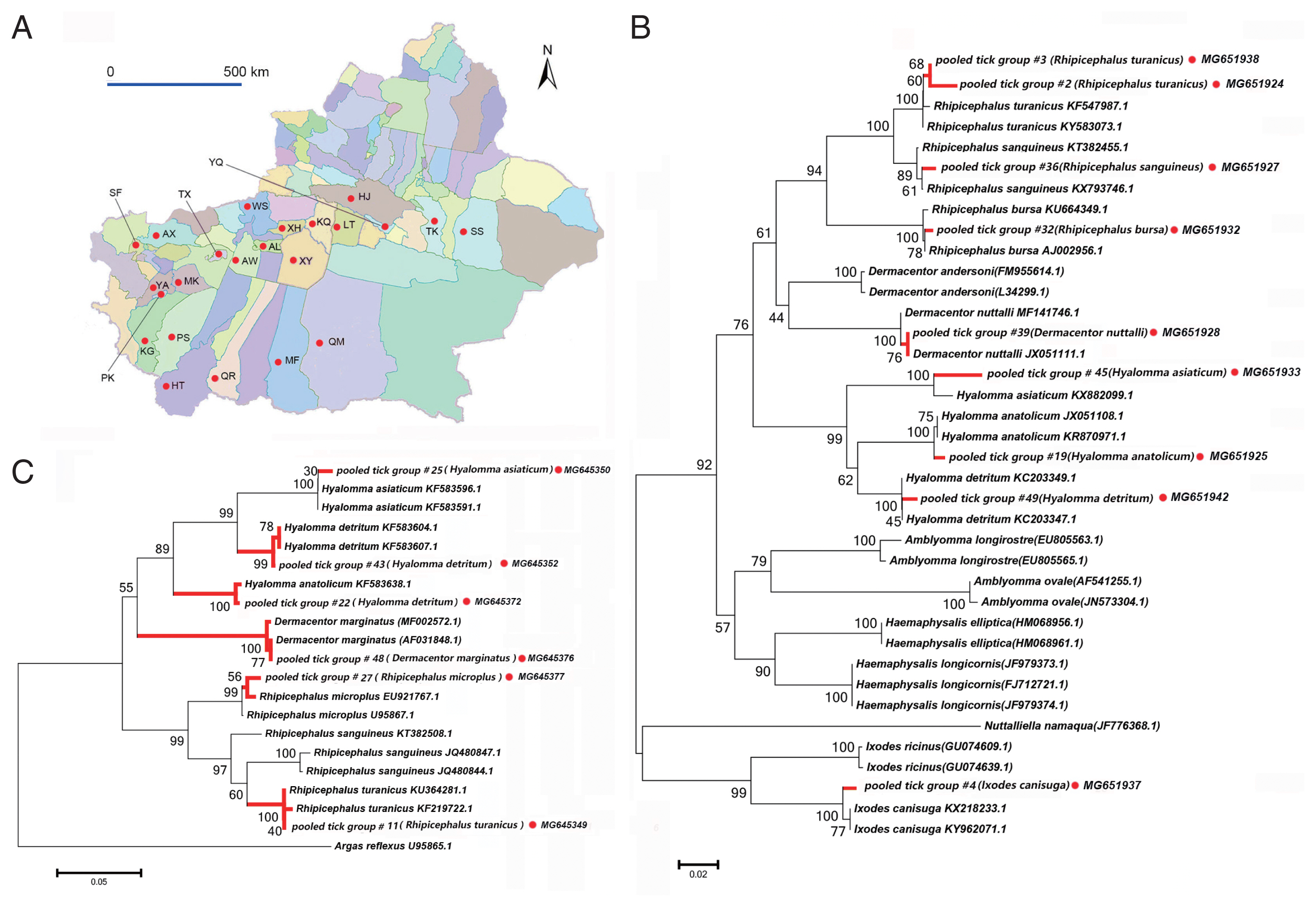
Xinjiang Uygur Autonomous Region is the largest province in China, covering more than 1/6 of the country’s territory; however, it is dominated by large areas of desert and mountain ranges [5]. Forty-two species of ticks from 9 genera have been identified in Xinjiang; these represent more than one-third of the total tick species identified in China [6]. More importantly, a high prevalence of tick-borne diseases has been reported in Xinjiang Autonomous Region [5]. The Tarim Basin is an endorheic basin in Xinjiang that covers an area of approximately 1,020,000 km2 [1]. The name Tarim Basin is sometimes used synonymously to refer to the southern half of the province, or southern Xinjiang. The northern boundary of the Tarim Basin is the Tian Shan mountain range and the southern boundary is the Kunlun Mountains along the edge of the Qinghai–Tibet Plateau [7]. Various landscapes of the Tarim Basin harbor of high mountains, valleys, plains, the Gobi Desert, and additional characteristics [8]. A previous survey of ticks from livestock conducted in northern Xinjiang indicated that the tick population was altered in response to climate change [5]. The climate of the Tarim Basin is drier than that of northern Xinjiang. The Tarim Basin is one of the most highly erodible areas in China, and the Taklimakan Desert covers 64% of this region [9]. Very low level of precipitation in the region inhibits plant growth and minimizes the habitable area for animals and humans [10]. In the Tarim Basin, 90% of the humans and animals inhabit oases that are supplied by run-off water from the surrounding mountains. This has the effect of concentrating humans and livestock around peripheral oases, the most important ecosystem in the area, which constitutes 4–5% of the total land cover of the Tarim Basin [11].
Until now, there have been few specific studies of ticks infesting domestic animals in these oases. However, little is known regarding tick-borne diseases of great medical significance, particularly zoonotic diseases, in this area. The current study investigated the associations between ticks and domestic animals in the Tarim Basin. In addition, molecular techniques were used to assess the presence of tick-borne pathogens.
MATERIALS AND METHODS
Sampling area and tick sampling and identification
From 2012 to 2016, we collected 3,916 ixodid tick specimens from 23 sampling sites in oases around the Tarim Basin, namely the Awat, Hejing, Hetian, Kargilik, Luntai, Minfeng, Poskam, Pishan, Qiemo, Qira, Shufu, Shanshan, Tuokexun, Xinhe, Xayar, Yarkand, Yanqi, Artux, Makit, Wensu, Tumxuk, Kuqa, and Aral counties (Fig. 1A). These counties are mainly regions of livestock husbandry. The livestock reared in these regions are primarily autochthonous. Most of the sheep (approximately 95%) are Kazakh, Chinese Merino, Hu, and Altay breeds. The cattle are mainly Xinjiang Brown, Holstein, and Kazakh breeds. The dogs are mainly Kazakh Tobet, a breed commonly used to protect livestock herds. During this 5-year survey, most sampling sites (17/23), including farms in the Awat, Hejing, Hetian, Kargilik, Luntai, Minfeng, Poskam, Pishan, Qiemo, Qira, Shufu, Shanshan, Tuokexun, Xinhe, Xayar, Yarkand, and Yanqi counties, were surveyed only once. Three sampling sites in the Artux, Makit, and Wensu counties were surveyed twice. The sampling site in the Tumxuk county was surveyed 3 times, and the farm in the Kuqa county was surveyed 5 times. The sheep flock on the sampling site in Aral county was surveyed twice, and the dogs were surveyed 6 times.
We collected adult ticks from the body surfaces of livestock and dogs over 5 consecutive years. Data recorded for all specimens collected included their location, host, number of ticks collected and the date of collection, species, and sex (Table 1). We grouped tick specimens according to location/host of collection, date of collection, and species. Ticks were collected from animals via tick sampling from the entire body of each animal, including sheep, cattle on 23 farms in 23 counties, and domestic dogs from farms in the Aral county. The collected ticks were kept alive until they were transferred to the laboratory of College of Animal Science, Tarim University. The collected ticks were counted and identified to species based on morphology according to standard morphological keys, and the identification was confirmed based on mitochondrial 16S and 12S rRNA sequences [12–14]. We utilized morphological characteristics, such as the anterior surface of the basis capituli flat, perpendicular to the longitudinal axis of the hypostome, for morphological species identification. All specimens were preserved in 70% ethanol. All livestock we checked for ticks were grazed in the traditional manner, and all farm dogs were free-roaming. There was no evidence of the use of chemical acaricides prior to tick sampling.
Nucleic acid extraction
We used 3 representative tick specimens from each specimen group for DNA extraction and mixed DNA templates of 3 ticks of each specimen group for molecular identification of tick species and screening of pathogens.
Ethanol-preserved tick samples were rinsed in distilled water, and then dried on filter paper. Total DNA was extracted using the DNeasy blood and tissue kit (Qiagen GmbH, Hilden, Germany) according to the manufacturer’s instructions. DNA was eluted in 80 μl elution buffer AE (provided with the kit) and stored at −80°C until tested.
Molecular identification of tick species
Partial sequences of 16S and 12S rRNAs were PCR amplified using the primers described in previous studies [14]. PCR amplification was performed using GoTaq G2 Flexi DNA polymerase (Promega Corporation, Madison, Wisconsin, USA). The reaction master mix was prepared according to the manufacture’s protocol, and PCR conditions described in previous studies were applied [15]. In brief, PCR reaction mixtures comprised 2 pmol of each primer, 200 μM of each deoxynucleoside triphosphate (dNTP), 1×PCR buffer, 1 unit of Taq DNA polymerase, and 50–100 ng of sample DNA for each PCR in a 50-μl reaction.
Total DNA (including mitochondrial) extracted from H. asiaticum was used as a positive control. Sterile water included in the DNA extraction procedure was used as a negative control. PCR products were electrophoresed on a 1% agarose gel, stained with SYBR Safe DNA Gel Stain (Invitrogen, Carlsbad, California, USA), and visualized under ultraviolet light. DNA bands of the correct size were purified and sequenced by BGI Tech Solutions Co., LTD (Liuhe, Beijing, China).
Sequences of 12S rRNA of ticks obtained in this study are deposited in GenBank under accession Nos. MG645336–MG645377. Sequences of 16S rRNA of ticks obtained in this study are deposited in GenBank under accession Nos. MG 651923–MG651949.
Detection of DNA sequences of tick-borne pathogens
A total of 156 specimens were screened for tick-borne pathogens. Piroplasms were detected using primers PIRO-A and PIRO-B, which produce 423-bp band [16]. Borrelia spp. were detected using a primer pair that targeted the flagellin gene [17]. Anaplasma spp. were detected using 2 primer pairs that targeted msp4 and 16S rRNA [14,18]. Rickettsia spp. was detected using a set of primer pairs targeting 6 DNA fragments, including partial sequences of 17-kDa antigen, 16S rRNA, citrate synthase (gltA), surface cell antigen 1 (sca1), outer membrane protein A (ompA), and outer membrane protein B (ompB), as described previously [19]. DNA bands of the correct size were purified and sequenced by BGI Tech Solutions Co., LTD. Representative DNA sequences of tick-borne pathogens are deposited in GenBank.
The reaction master mix was prepared according to the manufacture’s protocol, and PCR conditions described in previous studies were applied. All PCRs were run with positive and negative controls. Genomic DNA sequences of Babesia divergens, Rickettsia spp. [spotted fever group (SFG)], A. phagocytophilum, and Bo. garinii were used as positive controls. Sterile water included in the DNA extraction procedure was used as a negative control.
Sequence analysis
DNA sequences were assembled using Lasergene version 12.1 (DNASTAR) and edited in MEGA 5.0 [20,21]. Sequence alignments were performed using ClustalW within MEGA 5.0, using default parameters (open gap penalty=10.0, extend gap penalty=5.0) prior to checking via visual inspection. Genetic distances were calculated based on the K2P model for all pairwise comparisons in the matrix using MEGA5.0. Bootstrapping (1,000 replicates) was used to estimate variance. Pairwise deletion was used for gaps/missing data. Based on K2P distances, phylogenetic trees were constructed with the combined data sets of major tick genera and tick-borne pathogens using the neighbor-joining method, as described previously [22]. For gltA, msp4, and groEL analyses, the sites of codon positions were tested.
RESULTS
Ticks collected and morphological identification
We collected 3,916 ticks of 10 species belonging to 4 genera from 23 sampling sites in the Tarim Basin (Fig. 1A). These data are summarized in Tables 1 and 2. We confirmed species identity via phylogenetic analysis using 16S (Fig. 1B) and 12S rRNA (Fig. 1C) sequences. All specimens belonged to the family Ixodidae. The species collected most frequently were Rhipicephalus turanicus (n=2,224, 56.8%), Hyalomma anatolicum anatolicum (n=861, 22.1%), Rh. bursa (n=231, 5.9%), H. asiaticum asiaticum (n=205, 5.2%), Rh. sanguineus (n=144, 3.7%), H. detritum detritum (n=133, 3.4%), Dermacentor marginatus (n=53, 1.4%), D. nuttalli (n=37, 0.9%), Rh. microplus (n=23, 0.6%), and Ixodes canisuga (n=5, 0.1%), in that order. Notably, 5 I. canisuga females were collected from a farm dog in Xinjiang, supporting the existence of I. canisuga in Xinjiang, as reported in a previous study [23,24].
Rickettsia spp. Detectied
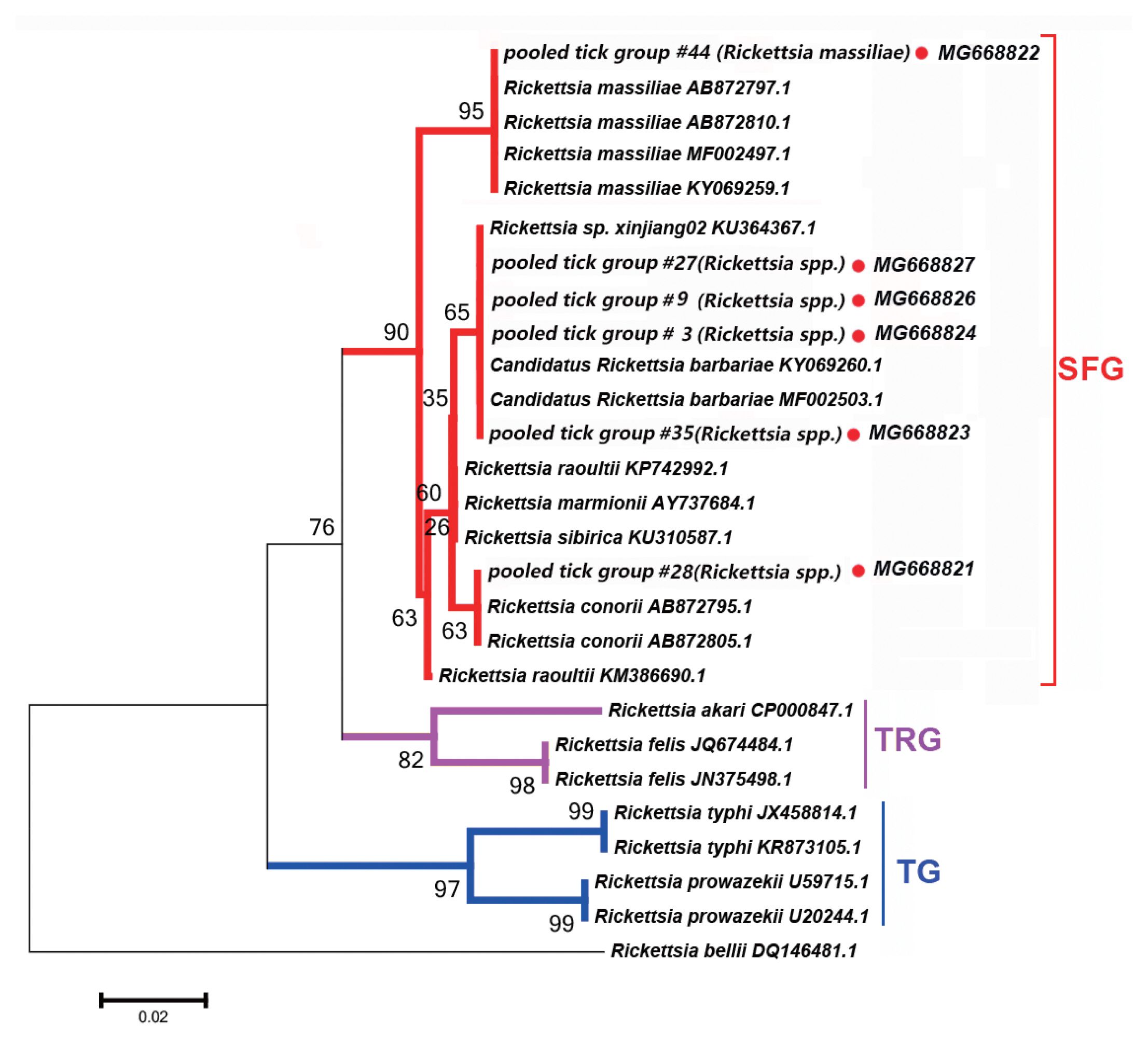
Six PCR products, namely 17-kDa antigen, gltA, sca1, 16S rRNA, ompA, and ompB, were assessed within each tick pooled tick group to examine the presence of Rickettsia spp. All PCR products of the correct size amplified from Rickettsia spp. were sequenced, and the obtained sequences of Rickettsia spp. obtained (Fig. 2).
We detected DNA sequences of Rickettsia spp. within SFG, such as Ri. conorii, Candidatus Rickettsia barbariae, and Ri. massiliae, in 11 of the 52 tick pooled tick groups (22.2%). This group included Rhipicephalus spp. collected from 8 counties in the Tarim Basin. Five of the 11 Rickettsia spp. of SFG were confirmed as Candidatus Ri. barbariae, one as Ri. massiliae, 1 as Ri. conorii, 1 as Ri. raoultii, 1 as Ri. sp. xinjiang02, and the remaining 2 as Rickettsia spp. without clear species identification. Fig. 2 depicts a phylogenetic tree based on the gltA sequences of the representative Rickettsia spp. Rickettsia sequences detected in this study were clustered with species within SFG, including Ri. conorii (AB872795.1) and Ri. massiliae (AB872810.1).
We detected Rickettsia spp. in Rh. turanicus, Rh. Bursa, and Rh. microplus ticks collected from the oases of the Makit, Tumxuk, Qiemo, Wensu, Qira, Aral, Minfeng, and Kuqa counties. We collected Rh. turanicus ticks from both sheep and cattle, Rh. bursa ticks from sheep, and Rh. microplus ticks from cattle.
Anaplasma spp. Detectied
We detected DNA sequences of Anaplasma spp. in 5 of the 52 tick pooled tick groups, which included Rh. turanicus and Rh. sanguineus ticks collected from the Aral and Kuqa counties. DNA sequence analyses of 4 amplicons from the Aral county revealed that they were identical to one another and shared 99% sequence identity with the 16S rRNA of Candidatus Anaplasma boleense (KX987335.1), whereas the amplicon from the Kuqa county shared 99% sequence identity with msp4 of A. ovis (KY283958.1). Fig. 3A depicts a phylogenetic tree based on the 16S rRNA sequences of the representative Ehrlichia/Anaplasma spp. The sequences acquired from the pooled tick groups #6 (Rh. turanicus), #7 (Rh. turanicus), #8 (Rh. sanguineous), and #9 (Rh. turanicus) belonged to the cluster corresponding to Candidatus A. boleense. Fig. 3B depicts another phylogenetic tree based on the msp4 sequences of the representative Anaplasma spp. In this analysis, the sequence acquired from pooled tick group #21 (Rh. turanicus) belonged to the cluster corresponding to A. ovis.
We detected Candidatus Anaplasma boleense in both Rh. turanicus and Rh. sanguineus. Rh. turanicus ticks were collected from both sheep and dogs, and Rh. sanguineus ticks were collected from dogs. We detected A. ovis by PCR in Rh. turanicus ticks, which were collected from cattle.
Borrelia spp. Detected
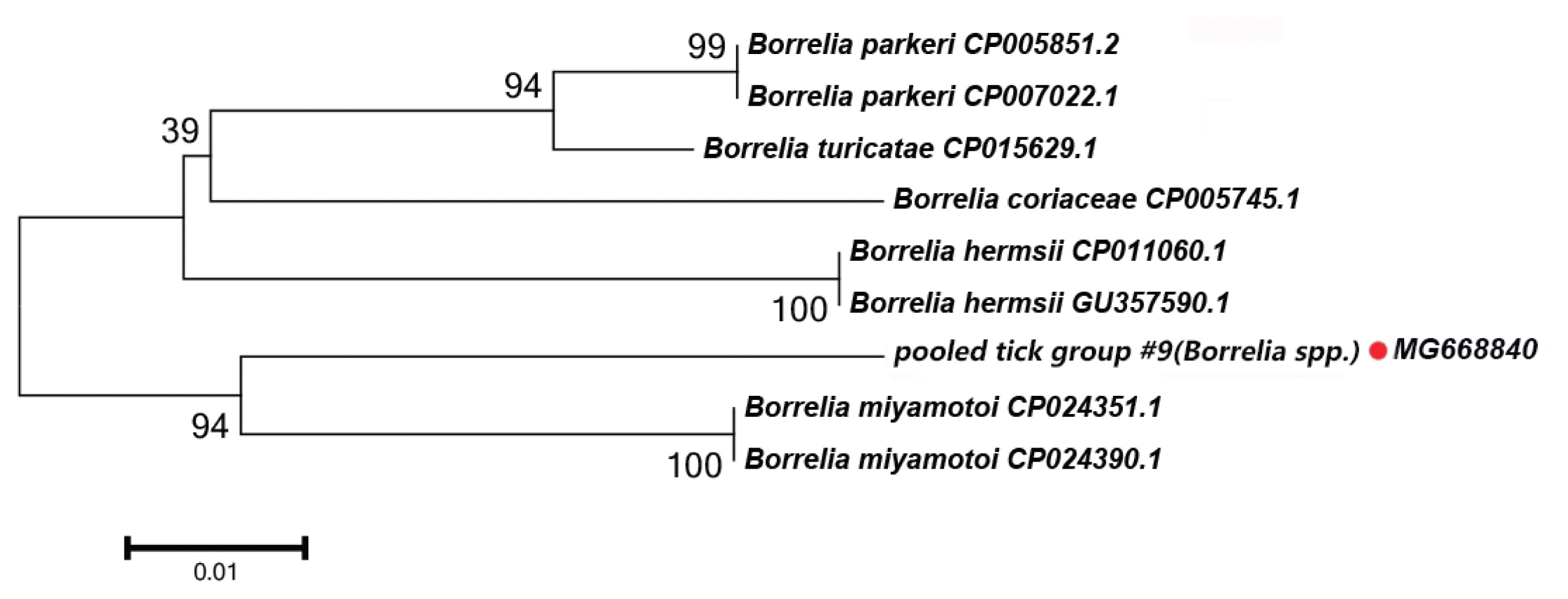
In this study, the groEL gene segment of Borrelia spp. was detected in Rh. turanicus collected from the Aral county; these ticks were collected from sheep. DNA sequence analysis of the amplicon revealed that it shared 94% sequence identity with the groEL gene of Bo. miyamotoi (CP004217.2). Fig. 4 depicts a phylogenetic tree based on the groEL sequences of the representative Borrelia spp. The DNA sequence of the groEL gene amplified in this study formed a unique cluster within Borrelia spp.
Babesia spp. Detected
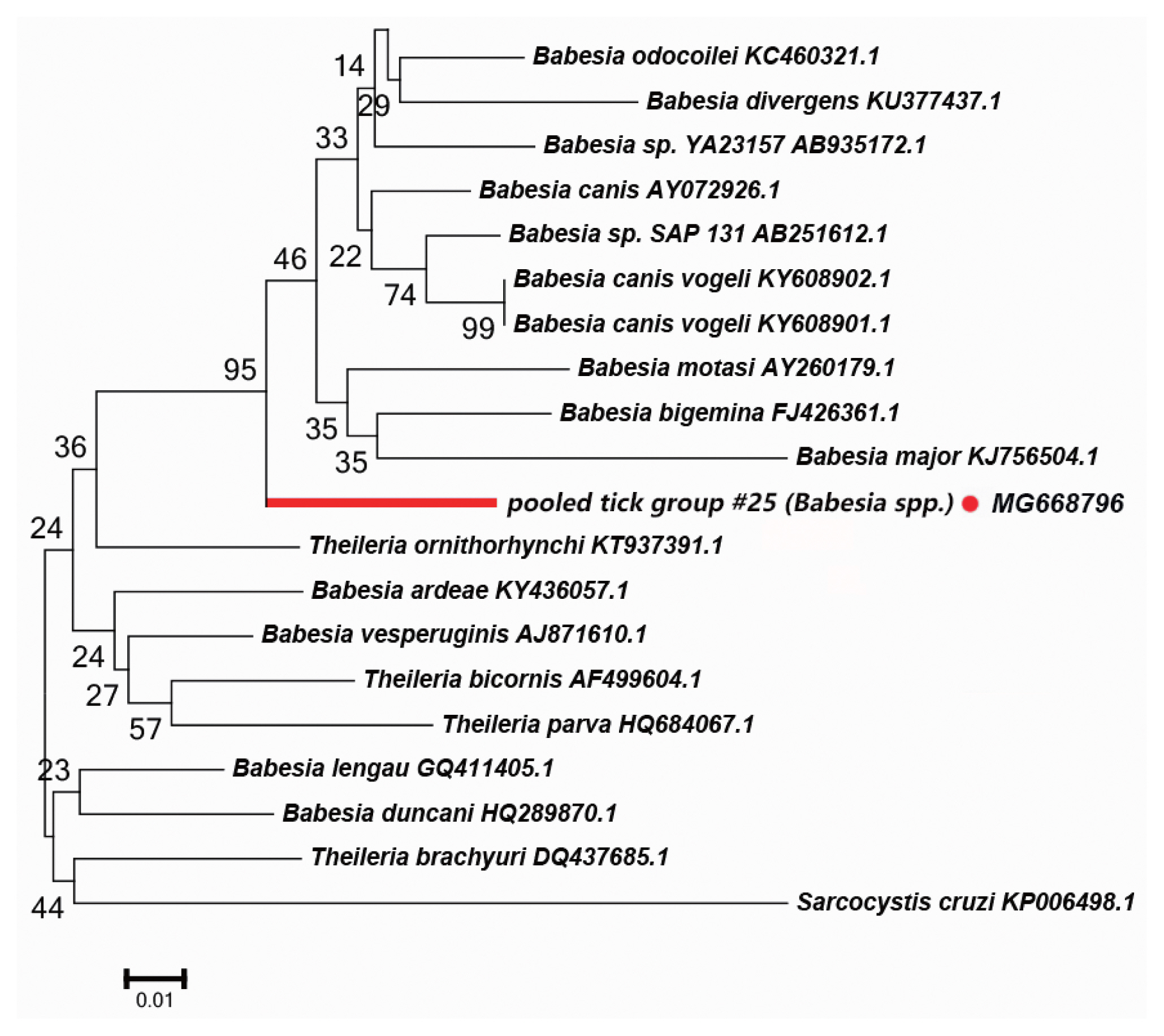
We detected a possibly undescribed Babesia spp. in H. asiaticum asiaticum collected from cattle in the Kuqa county. DNA sequence analysis of the amplicon revealed that it shared 91% sequence identity with the 18S rRNA sequence of a Ba. canis vogeli isolate (KT323935.1). Fig. 5 depicts a phylogenetic tree based on the 18S rRNA sequences of the representative Babesia/Theileria spp. The DNA sequence of 18S rRNA amplified in this study formed a unique cluster within Babesia/Theileria spp.
DISCUSSION
The survival, phenology, and biting activities of ticks are closely reliant on environmental conditions, including annual rainfall, atmospheric temperature and relative humidity, vegetation cover, altitude, and host availability [5]. There is a dearth of study of disease transmission by ticks, and such studies necessitate ongoing active surveillance of the distribution of ticks in areas with specific desert environments, such as the Tarim Basin.
We identified 10 tick species from 4 genera associated with domestic animals from 23 study sites (Fig. 1A). These species included Rh. turanicus, H. anatolicum anatolicum, Rh. bursa, H. asiaticum asiaticum, Rh. sanguineus, H. detritum detritum, D. marginatus, D. nuttalli, Rh. microplus, and I. canisuga, most of which have been reported in the study area. However, our findings are not entirely consistent with those of previous studies [3]. We found Rh. turanicus to be the most widespread tick species, although previous studies have described Rh. turanicus as the second most abundant tick species in southern Xinjiang [25]. We collected Rh. turanicus from domestic animals in oases in 14 counties. Rh. turanicus constituted 56.79% of the total number of ticks collected (2,224/3,916), and it was highly prevalent in various domestic hosts, including cattle, sheep, and dogs. Rh. turanicus has been implicated as a vector of several human and veterinary pathogens, including Rickettsia spp. of SFG, Anaplasma spp., Borrelia spp. and West Nile virus [26]. Rh. turanicus can infest people, and the risk of pathogens being transmitted from Rh. turanicus to people should not be ignored [27].
In Xinjiang, I. canisuga was collected from a dog that had never traveled outside the region. We confirmed the morphological identification of I. canisuga using molecular and phylogenetic methods. DNA sequence analysis of the amplicon revealed that the 16S rRNA sequences of the pooled tick group #4 shared 98.47% sequence identity with the 16S rRNA sequences of I. canisuga from Hungary (KX218233.1). Precise distribution of I. canisuga in Xinjiang requires further study. To date, I. canisuga has been recorded in Xinjiang just once [24]. There are several potential reasons for this. First, I. canisuga is a common ectoparasite of carnivores; however, previous surveys in Xinjiang have always collected ticks from livestock and not from carnivores [28]. Second, previous studies have revealed the proportion of I. canisuga in some areas to be relatively rare [29]. We identified 3 tick species from dogs and found that the proportion of I. canisuga in these ticks was 4.42% (5/113). Our findings support previous reports of the presence of I. canisuga in Xinjiang [24].
In this study, we detected several pathogens in ticks from southern Xinjiang, including the Rickettsia spp. of SFG and Anaplasma spp. To screen for the Rickettsia spp., we assessed 6 partial sequences of 17-kDa antigen, 16S rRNA, gltA, sca1, ompA, and ompB. Coinfections of multiple species of Rickettsia in a single tick may occur [30]. Tick-borne pathogens such as Candidatus Anaplasma boleense, Candidatus Rickettsia barbariae, and Ri. massiliae have been reported in China [31,32], which were also detected in this study. However, it is noteworthy that possibly undescribed Borrelia and Babesia spp. were detected in this study.
In Borrelia spp., the distribution of inter-specific K2P distances for groEL at the species level was between 1.8% and 9.7%. The groEL DNA segment that we detected could be correctly translated into a protein according to the Bacterial, Archaeal, and Plant Plastid Code, and it is nested within a novel phylogenetic group in the Borrelia spp. clade (Fig. 4). We speculate that the groEL DNA segment detected in this study belongs to an undescribed Borrelia sp.; multilocus gene typing by targeting other genes should be performed.
In the Babesia spp., the distribution of inter-specific K2P distances for 18S rRNA at the species level was between 0.8% and 14.4%. The 18S rRNA we detected was nested within in a novel phylogenetic group in the Babesia spp. clade (Fig. 5). Molecular evidence from recent studies has revealed the presence of some new Babesia sp. [33]. We speculate that the 18S rRNA segment we detected in this study belongs to an undescribed Babesia sp; further study is required to confirm this.
In this study, 3,916 ixodid ticks (Acari: Ixodidae) from 10 species of 4 genera, which infest domestic animals, were collected from peripheral oases in the Tarim Basin. The study revealed that Rh. turanicus is currently the dominant ixodid species in this area. We rediscovered I. canisuga from Xinjiang after its first record 45 years ago. In addition to the tick-associated microorganisms, we detected some potential zoonotic pathogens, including Rickettsia spp. of SFG and Anaplasma spp. The analyses suggested the presence of some hitherto undescribed Babesia spp. and Borrelia spp. in ticks in the Tarim Basin. Some of the tick species we collected, such as Rh. turanicus and Rh. Sanguineus, may infest people, and they represent a potential risk of disease transmission from ticks to both livestock and people in the Tarim Basin.