INTRODUCTION
Giardia lamblia, a protozoan pathogen which causes a diarrheal disease in man, has a simple life cycle that is composed of 2 forms, trophozoites and cysts. Upon ingestion of the cysts, its infection is initiated and this is followed by excystation to the trophozoite form usually in the proximal duodenum of the host. Trophozoites are able to multiply by binary fission and colonize the proximal small intestine using adhesive discs. Some trophozoites undergo encystation, i.e., differentiation of trophozoites to the cyst form, in the distal small intestine. The distinct structural differences between these 2 forms imply that a series of genes are differentially expressed during these differentiations.
Previous studies on the encystation of G. lamblia identified several encystation-induced genes such as cwp1, cwp2 (Mowatt et al., 1995), bip/grp78 (Lujan et al., 1996), and enc1 and enc6 (Que et al., 1996). Knodler et al. (1999) reported that the expression of gln6pi-b for glucose-6-phosphate isomerase was also induced during encystation. In addition, the expression of the myb2 gene encoding a putative transcriptional factor was found to be induced during encystation (Sun et al., 2002; Yang et al., 2003). Investigation of the 2 gap genes, gap1 and gap2 of G. lamblia (Rozario et al., 1996) indicated that Gap1 protein functions as a major glycolytic enzyme whereas gap2 does not seem to encode a protein with GAPDH activity (Yang et al., 2002). Despite the isolation of the gap2 gene as an induced clone in a differential display of encysting cells of G. lamblia, no detailed examination has been performed on its expression and function. In the present study, the expression of the gap2 gene was monitored using transgenic G. lamblia carrying a gap2-luc reporter system.
MATERIALS AND METHODS
Organisms and culture conditions
Trophozoites of G. lamblia WB strain (ATCC 30957, Washington DC., U.S.A.) were grown for 72 hr in normal TYI-S-33 medium (2% casein digest, 1% yeast extract, 1% glucose, 0.2% NaCl, 0.2% L-cysteine, 0.02% ascorbic acid, 0.2% K2HPO4, 0.06% KH2PO4 and 10% calf serum) (Keister, 1983) containing 0.5 mg/ml bovine bile at pH 7.1, and then transferred into an encystation medium containing 10 mg/ml of bovine bile at pH 7.8 (Kane et al., 1991). Portions of the encysting cells were sampled at various times by chilling on ice and centrifugation.
Construction of plasmids containing a luc reporter gene
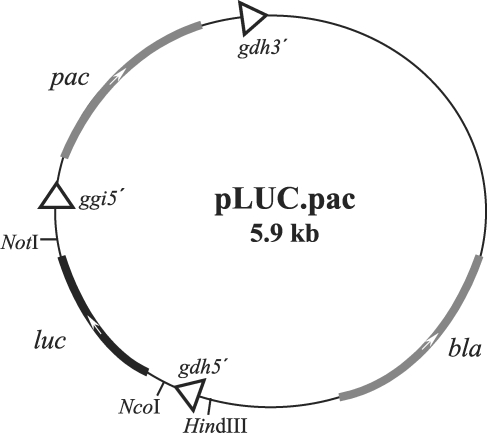
To obtain a reporter system to allow us quantitative measurement of activity of the gap2 gene promoter in G. lamblia, we made a new vector based on pGFP.pac (Singer et al., 1998). A 1,682-bp DNA fragment containing the full ORF of the luc gene was amplified from pGL2-Basic vector (Promega, Madison, U.S.A.). by PCR using 2 primers, LUC-F and LUC-R (Table 1). An NcoI and a NotI site located at either ends of the resultant luc DNA were used to clone this DNA into the corresponding sites of pGFP.pac, which resulted in plasmid pLUC.pac, in which the gfp gene was replaced by the luc gene (Fig. 1).
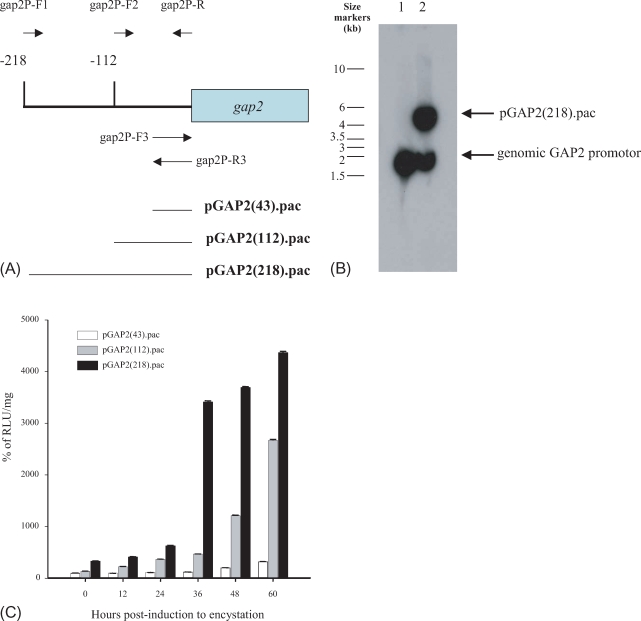
Based on the sequence published by Rozario et al. (1996) (GenBank database accession number, U31911), 3 different DNA regions of the gap2 promoter were amplified and cloned into pLUC.pac (Fig. 2A). First a 218-bp region of the DNA upstream of the gap2 gene was amplified from the genomic DNA of G. lamblia using 2 primers, gap2P-R and gap2P-F1 (Table 1), and inserted into pLUC.pac pretreated with HindIII and NcoI, which resulted in a deletion of the gdh promoter. The second promoter studied was a 112-bp DNA upstream region of the gap2 gene, which was made from the genomic DNA of G. lamblia with the primers, gap2P-R and gap2P-F2. The third gap2-luc reporter containing a 43-bp upstream region of ATG of the gap2 ORF was constructed by cloning the annealed linker of 2 primers, gap2P-F3 and gap2P-R3, into the HindIII and NcoI site of pLUC.pac.
Transfection
Trophozoites were grown for 72 hr in normal TYI-S-33 medium. Fifteen µg of a gap2-luc reporter plasmid
was transformed into 1 × 107 trophozoites by electroporation
under the following conditions; 350 volts, 1,000 µF, and 700 Ω (Biorad Genepulser II, Hercules, U.S.A.). Trophozoites harboring a gap2-luc plasmid were selected by adding puromycin to the TYI-S-33 medium to a final concentration of 100 µM.
Determination of luciferase activities
Luciferase activities of the harvested cells carrying one of the gap2-luc reporter plasmids were measured using a Luciferase Assay System (Promega). Briefly, collected cells were resuspended in a lysis buffer (25 mM Tris-HCl, pH 7.8, 2 mM EDTA, 2 mM DTT, 10% glycerol, and 1% Triton X-100), and frozen at -70℃ at least for 1 hr Twenty µl of cell extracts were reacted with 100 µl of luciferase substrate, and light emission was measured for 5 min in a luminometer (TD20/20 DLReady, Turner Designs, Sunnyvale, U.S.A.).
Southern blot analysis
Genomic DNAs were purified from untransfected trophozoites as well as from trophozoites transfected with pGAP2(218).pac. Ten µg of genomic DNA was digested with HindIII and loaded into a 0.8% agarose gel. Upon separation by electrophoresis, digested genomic DNA was transferred to a nytran filter (Millipore, Billerica, U.S.A.), and fixed to the filter by UV-crosslinking (Hoefer, San Francisco, U.S.A.). The filter was then hybridized using the 218-bp gap2 promoter region, which was labeled with 32P by using a Random labeling kit (Takara, Otsu, Japan).
RESULTS
Construction of 3 gap2-luc reporter systems
We used the luciferase reporter system to examine the expression of the gap2 gene, which has been identified to be induced during encystation (Yang et al., 2002). To define the essential cis-acting elements required for the full expression of the gap2 gene, 3 different gap2-luc fusions were constructed, which contained gap2 gene promoter regions of different sizes, as described in Fig. 2A.
Confirmation of transgenic G. lamblia containing a gap2-luc reporter plasmid
All of the 3 gap2-luc constructs were transfected into trophozoites as described above. For each construct, 2 independent transfectants were selected, and used for further studies. Maintenance of the gap2-luc reporter in puromycin-resistant G. lamblia was verified by Southern blot analysis (Fig. 2B). Genomic DNA of untransfected trophozoites displayed a band of ~1.7 kb in Southern blots with a gap2 promoter region. In the case of genomic DNA of trophozoites transfected with pGAP2(218).pac, an additional band of 5.8 kb DNA was found in addition to the ~1.7 kb DNA band, which demonstrated that pGAP2(218).pac was maintained in puromycin-resistant G. lamblia clones.
Determination of the expression of the gap2 gene in G. lamblia
For each gap2-luc construct, 2 independent transfectants were selected and their luciferase activities were measured. After establishing them as stable clones, the transfectants were induced to encystation by cultivating them in TYI-S-33 medium with high bovine bile at alkaline pH. At 5 different time-points of postencystation, i.e, 12, 24, 36, 48, and 60 hr, portions of encysting cells were harvested.
In the trophozoite form, the 3 gap2-luc fusions displayed differential luciferase activities (Fig. 2C). Transfectants carrying pGAP2(43).pac showed lower luciferase activities than those of trophozoites carrying 1 of the other 2 constructs, pGAP2(112).pac and pGAP2(218).pac. Trophozoites of G. lamblia carrying pGAP2(112).pac showed 40% more luciferase activity than those carrying pGAP2(43).pac. Trophozoites carrying pGAP2(218).pac showed the highest luciferase activity, namely, over 3 fold higher than that of Giardia trophozoites containing pGAP2(43).pac.
Upon encystation, Giardia containing these 3 constructs showed significant increases in luciferase activity, even though the fold increases were different for each construct. Transfectants carrying pGAP2 (43).pac, also showed higher luciferase activities during encystation than that of trophozoites, i.e., gradual increase of up to 3-fold. Giardia containing pGAP2 (112).pac also showed a gradual increase in luciferase activity as cells proceeded to encystation (20-fold). G. lamblia carrying pGAP2(218).pac also displayed a dramatic increase in luciferase activity upon encystations; a 13-fold increase at 60 hr post-induction.
DISCUSSION
Despite the putative role of glyceraldehydes 3-phosphate dehydrogenase implied from the amino acid sequences of the gap2 ORF, no enzymatic activity was demonstrated in the previous study (Yang et al., 2002). To identify the role of this putative gap gene, we confirmed encystation-induced expression of the gap2 using a luciferase reporter system. There is no information on the cis-acting elements required for transcription in G. lamblia at the present level of knowledge. Therefore, we randomly constructed 3 luc fusions containing different sizes of the gap2 promoter, i.e., 43, 112 and 218-bps.
When the constructed plasmids were transfected into G. lamblia, we found that the efficiency of transfection was very low, i.e., below 10-7. Therefore, an experiment to detect the presence of the gap2-luc reporter in tranfected G. lamblia was performed to exclude a possibility that we could select puromycin-resistant clones of G. lamblia during transfection (Fig. 2B).
Luciferase activities of 3 different gap2-luc fusions clearly showed that 43-bp upstream region of the initiation codon of the gap2 ORF did not contain all the essential elements required for expression of the gap2 gene (Fig. 2C). In addition, the 112-bp upstream region of the gap2 gene seemed to have cis-acting information for the full induction of this gene during encystation.
In this study, we constructed a reporter system in G. lamblia, which allows the quantitative measurement of the expression of a prescribed gene. This system was applied to monitor the expression of gap2, which was identified as one of the genes induced during encystation. Using reporter constructs containing different regions of the gap2 promoter, we defined the cis-acting elements required for full induction of the gap2 gene during encystation.