INTRODUCTION
Since 1993, when an indigenous case of Plasmodium vivax infection was reported (Chai et al., 1994), more than 10,000 cases of vivax malaria have occurred in the south-west and near the demilitarized zone (DMZ) of Korea (Feighner et al., 1998; Lee et al., 1998; reviewed by Chai, 1999; Ree, 2000). The reemergence of vivax malaria has been presumed to expand from the endemic regions in the north of DMZ mainly by the changes in the vector environments, although there are no information available on the endemic status in the north. Unique clinical features of the prolonged incubation period and genetic approaches (Kho et al., 1999; Lim et al., 2000) demonstrated that the prevalent strain had very similar characteristics to the North Korean strain described by Shute et al. (1977).
Microscopic examinations of Giemsa-stained thick and thin blood smears (BS) have been the diagnostic method of choice (Warhurst and Williams, 1996). However, there are two limitations when detecting vivax malaria: one is caused by the biology of vivax malaria and the other by the examiner. It is not possible to observe parasites by BS during the irregular prolonged incubation periods of vivax malaria in the temperate climate regions (Krotoski, 1985) of which the incubation periods vary from 153 to 452 days before the onset of malarial symptoms in the Korean cases (Lee et al., 1998). And the other limitation of BS includes the lack of well-trained personnel and the length of time required for the examination, especially when parasitemia is as low as those in P. vivax infections.
Various detection methods have been developed to overcome these limitations such as antigen- (Shiff et al., 1993; Dietze et al., 1995) and nucleic acid-based detections (Barker et al., 1992; Li et al., 1995) of falciparum malaria. Antibody-based detection methods such as indirect haemagglutination test (WHO, 1988), indirect fluorescent antibody test (Mendis et al., 1987), and ELISA tests (Demedts et al., 1987; Del Giudice et al., 1987) have also been established. Up until now, western blot (WB) has not been performed as a method of serological diagnosis of malaria. In an attempt to establish a WB diagnosis of vivax malaria, we carried out WB with patients' sera against multiple recombinant antigens selected as stage-specific antigens to vivax malaria.
MATERIALS AND METHODS
Examination of blood smear and plasma collection
Thin blood films were stained with Diff-Quick solution (International Reagents Corp., Kobe, Japan) and examined under oil-immersion (100X) for 10 fields. Plasma was collected after centrifugation of the whole blood at 12,000 rpm and frozen at -70℃ until use. With this method, 160 cases of P. vivax infections were diagnosed. And 13 additional cases of P. falciparum infections and 9 additional cases of P. vivax/P. falciparum mixed infections from endemic African or southeast Asian nations were also evaluated.
Polymerase chain reaction (PCR)
The DNA was extracted from the whole blood (200 µl) of a vivax malaria patient using a QIAamp DNA mini kit (QIAGEN, Valencia, CA) according to the manufacturer's protocol. Primers were synthesized as in Table 1 on the coding regions for the antigenic domains of circumsporozoite protein (CSP-1, GeneBank accession No. M34697), merozoite surface protein (MSP-1, M60807), apical merozoite antigen (AMA-1, AF063138), serine repeat antigen (SERA, AF052747), and exported antigen (EXP-1, X05074) of Plasmodium vivax. Denaturing and extension temperatures used for PCR were 95℃ for 1 min and 72℃ for 2 min for 30 cycles. The annealing temperature of each reaction was described in Table 1.
Production of recombinant GST fusion antigens
The amplified DNA fragments of CSP-1, MSP-1, AMA-1, SERA, and EXP-1 were cloned into pGEM-T Easy vector (Promega, Madison, WI) to subclone in pGEX-4T vector (Amersham Pharmacia Biotech, Uppsala, Sweden). An inframe insertion of each plasmid was confirmed by sequencing the vectors with T7 and SP6 sequencing primers. Each protein was expressed as GST fusion proteins in Escherichia coli (HB101 strain) by the isopropyl-β-D-thiogalactoside (IPTG) induction for 3 hr at 30℃.
Western blot
Western blot was performed by the method of Towbin et al. (1979). The whole bacterial cell extracts were separated in 10% SDS-PAGE gels and transferred onto nitrocellulose sheets (NC, Schlleicher and Shuell, Keene, NH). NC membrane, blocked with 5% skim milk in PBS/0.05% Tween-20, was incubated with serum of 1:500 dilution followed by a subsequent incubation with 1:2,000 diluted HRP-conjugated goat anti-human IgG antibody (Cappel, Costa Mesa, CA). The membrane was soaked in enhanced chemiluminescence (ECL) solution (Intron, Daejon, Korea) for 1 min and exposed to an X-ray film (Konica, Tokyo, Japan). Positive reaction was determined by at least one or more bindings with specific antigens, and the arbitrary band intensity was measured after scanning the films.
RESULTS
Amplification of gene fragments by PCR and expression as GST fusion proteins
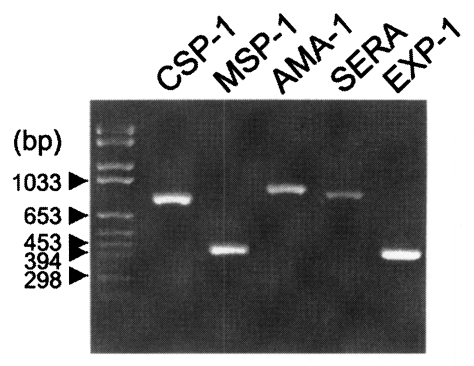
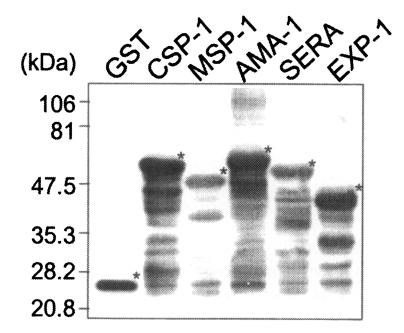
The coding regions for the antigenic domains of CSP-1, MSP-1, AMA-1, SERA, and EXP-1 of P. vivax were amplified by PCR as 774, 456, 894, 867, and 423 bp as shown in Fig. 1. Amplified DNA fragments were cloned into pGEM-T Easy vector to subclone in pGEX-4T vector with the restriction of Eco RI. In-frame insertion of each plasmid was confirmed by sequencing the vector with T7 and SP6 sequencing primers. Each protein was expressed as the GST fusion protein in HB101 bacteria by the IPTG induction with the molecular mass of 58 kDa for CSP-1, 48 kDa for MSP-1, 60 kDa for AMA-1, 56 kDa for SERA, and 42 kDa for EXP-1, respectively, by the GST detection as marked with asterisks (*) in Fig. 2.
Detection of GST fusion antigens by patients' sera
All GST fusion proteins were antigenic sufficiently against sera of malaria patients without binding to GST only. When applied with patients' sera, 147 (91.9%) out of 160 vivax malaria, 12 (92.3%) out of 13 falciparum malaria, and all 9 vivax/falciparum mixed malaria reacted with at least one antigen, while no reactions were observed with 20 normal uninfected sera (Table 2).
In cases of vivax malaria, CSP-1 reacted with 128 (80.0%) sera, MSP-1 with 102 (63.8%), AMA-1 with 128 (80.0%), SERA with 115 (71.9%), and EXP-1 with 89 (55.6%), respectively. Among the 147 vivax malaria positive reactions, 10 sera (6.8%) reacted with only one antigen, 15 sera (10.2%) with two antigens, 22 sera (15.0%) with three antigens, 43 sera (29.3%) with four antigens, and 57 sera (38.8%) with all five antigens used. As shown in Table 3, higher detection rates were obtained when using 5 antigens (91.9%) rather than using each antigen solely (55.6 - 80%), a combination of 2 (76.3 - 87.5%), 3 (85.6 - 90.6%), or 4 antigens (89.4 - 91.3%).
And in cases of falciparum and vivax/falciparum mixed malaria, EXP-1 reacted strongly with sera in all western blot positives.
DISCUSSION
The GST fusion recombinant proteins coding the antigenic domains of CSP-1, MSP-1, AMA-1, SERA, and EXP-1 of P. vivax were analyzed in western blot with patients' sera to establish a diagnostic method. Each antigen was highly specific and reactive to some sera but the sensitivity was not enough when used solely, which resulted from the complexity in humoral immunity of vivax malaria (Krotoski, 1985). However, when four or five antigens were combined as detection antigens, positive reactions were identified as above 90%. There is space to be improved, this western blot with multiple antigens can be used as an antibody-based detection method.
Even though all samples were confirmed to be infected with the parasite by blood smear examination, sera of 13 cases (8.1%) in 160 vivax malaria and 1 case (among 13 cases) of falciparum malaria did not react in western blot. These vivax cases were presumed to be immediate onsets according to the due course without prolonged pre-erythrocytic stage, which might occur before the production of antibodies to the parasite.
And most imported malaria cases were caused by P. falciparum infection in Korea. Most of the travelers to the tropic and subtropic endemic regions have been guided to have prophylaxis against malaria, which results in the prevention in most travelers regardless of mosquito biting. But in some cases, not protected completely, malarial symptoms were suppressed merely during stay in the endemic region. Antibodies preformed before the onset of symptoms by recrudescence were detected in western blot in most imported cases except for 1 case in this study.
Western blot diagnosis with circumsporozoite and hypnozoite stage antigens will play critical role in the diagnosis of prolonged incubation period, especially in vivax malaria. And with merozoite antigens in addition to the antigens of exo-erythrocytic stages may provide useful information on the clinical procedure of vivax malaria. It can also be used in the epidemiological screening and safety detection of blood bank donors. Army soldiers have served as a large pool of blood donors in Korea. But the overlap of endemic regions and garrison camps (Kho et al., 1999) makes the blood supply to be perilous. Repeated mass screening methods may be helpful to select healthy donors under this situation, we propose a seasonal screening during winter when the mosquitoes not active.