INTRODUCTION
Intestinal mastocytosis is observed in certain intestinal helminth infections. The increase of mast cell in the intestinal mucosa is known to play an important role in host defense against intestinal parasites (Miller, 1984). At least three gastrointestinal nematode parasites have been identified: Trichinella spiralis, Strongyloides ratti, and Strongyloides venezuelensis (Ha et al., 1983; Abe et al., 1992; Grencis et al., 1993; Khan et al., 1993; Donaldson et al., 1996; Lantz et al., 1998). The expulsion of these parasites is severely impaired in mast cell-deficient W/Wv mice and in mice deficient in the mast cell-stimulating cytokine, IL-3 (Lantz et al., 1998). Intestinal mastocytosis is also observed in rats infected with tapeworms such as Hymenolepis diminuta and H. microstoma and intestinal trematode such as Fibricola seoulensis (Andreassen et al., 1978; Novak and Nombrado, 1988; Kho et al., 1990). However, the role of mucosal mast cell (MMC) has not been completely elucidated and there is lack of information on MMC responses against intestinal trematode infection.
Echinostoma hortense was first discovered by Asada (1926). In Korea, the human cases have been increased gradually since the first case of human infection was reported in the early 1980s (Seo et al., 1983). In experimentally rats infected with E. hortense, the worms are found in the intestinal villi from day 1 to day 3 p.i. and in the lumen from day 7 to day 44 p.i.. Intestinal infection causes villous loss, villous atrophy, and crypt hyperplasia (Lee et al., 1990). E. hortense is primarily located in the duodenum and the jejunum of infected rats.
In the present study, we observed the kinetics of intestinal MMCs at various sites of the small intestine of rats infected with E. hortense and compared it to the worm expulsion.
MATERIALS AND METHODS
Experimental infection and worm recovery
Metacercariae of E. hortense were isolated from the muscles of raw loaches, Misgurnus anguillicaudatus, obtained from Sumgin-gang (River), Kurye, Chonnam, Korea, by using artificial gastric juice. Sprague-Dawley rats weighing approximately 150 gm were orally infected with 150 metacercariae through a polyethylene capillary tube. The rats were sacraficed in each week p.i. and the worms were collected from the small intestines of the rats.
Observation of MMC responses
MMC was examined in the small intestines of rats on day 3, 5 and each week for day 56 p.i.. For histological samples, the intestinal segments were cut approximately 1.5-2 cm each in length and were taken from three sites of the small intestine: the duodenum (10 cm posterior to the pylorus), and the upper and lower parts of the jejunum. The excised segments of the small intestine were fixed in Carnoy's solution for 2-4 hrs. The fixed tissues were embedded in paraffin and sectioned at about 5 µm thickness using microtome. The sectioned samples were stained with 1% alcian blue (pH 0.3) and counter-stained with 0.5% safranin (pH 1.0) (Strobel et al., 1981). Stained MMCs were counted in the graticule (500 × 500 µm). The values of of MMC were recorded as the average number of MMC per graticule (0.25 mm2).
RESULTS
Kinetics of MMC responses
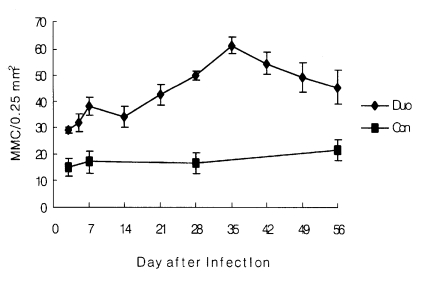
In the experimentally infected group, the number of MMC in the duodenum increased slightly from day 3 (29.0 ± 1.0) to day 5 p.i. (31.7 ± 3.4). At 14 day p.i., it decreased slightly but began to increase from day 21 p.i. It peaked on day 35 p.i. (61.3 ± 3.1); thereafter, it decreased slowly until day 56 p.i. (Table 1, Fig 1 and 2). The number of MMC in the upper and the lower parts of the jejunum was relatively lower than that in the duodenum. However, the kinetics of MMC responses were similar (Table 1).
In contrast, no evident changes in the kinetics of MMC responses was shown in non-infected control group (Table 1).
Worm recovery rate
The worm recovery rate decreased slowly from day 7 to 35 p.i.. It continued to decrease apparently from day 42 to day 56 p.i., 20.8% on day 42, 12.9% on day 49 and 5.5% on day 56 p.i. (Table 2).
DISCUSSION
This study shows that intestinal mastocytosis was induced in rats infected with E. hortence and that this response may be associated with explusion of the worm. The highest number of MMC was observed in the duodenum where the most of the worms dwelled. Similar result about the mast cell response was shown in the small intestine of rats infected with S. ratti (Mimori et al, 1982) In contrast to our results, Kho et al. (1990) reported that the increase in MMC number of rats was limited to the upper part of the small intestine in F. seoulensis infection. In this experiment, the peak level of mastocytosis was observed at day 35 p.i.. the value decreased thereafter. The worm recovery rate was evidently decreased on day 49 and day 56 when the higher level of mastocytosis was persisted. In contrast to our results, the peak level of mastocytosis was observed after the worms were expelled in T. spiralis infection or S. ratti infection (Woodbury et al., 1984; Mimori et al., 1982). In F. seoulensis infection, however, the peak level of mastocytosis was observed on the same time when the worm recovery rate began to decrease significantly. These different aspects between their and our results may be due to the different biological properties of the species of parasites.
There are many suggestions that MMC may play an important role in the host defense against intestinal parasites. Recent studies, however, showed that the effector cell in immune response against the parasites is not mast cell but goblet cell. It was reported, using concurrent infection with S. ratti and N. brasiliensis in rats, that mast cells were important in the expulsion of the former but not of the latter (Nawa and Korenaga, 1983). In mice infected with E. trivolvis, it was observed that the expulsion was associated mainly with goblet cell hyperplasia (Weinstein and Fried, 1991; Fujino et al., 1993). Fujino et. al. (1996) re-ported that goblet cell hyperplasia was inhibited and the number of mast cells and eosinophils increased when the expulsion of E. trivolvis was delayed by dexamethasone treatment. Considering the above these studies, main effector cells of immune responses in the intestinal paraisite infection may be different according to the species of intestinal helminths and kinds of hosts.
In conclusion, we suggest that MMC in rats are associated with the expulsion of E. hortense. In order to reach a more definite role of mast cells in protective immunity to E. hortense infection, we will need to further study on other effector cells such as goblet cells.