Warning: mkdir(): Permission denied in /home/virtual/lib/view_data.php on line 81
Warning: fopen(upload/ip_log/ip_log_2024-05.txt): failed to open stream: No such file or directory in /home/virtual/lib/view_data.php on line 83
Warning: fwrite() expects parameter 1 to be resource, boolean given in /home/virtual/lib/view_data.php on line 84 Development of Eimeria tenella in MDBK cell culture with a note on enhancing effect of preincubation with chicken spleen cells
Development of Eimeria tenella in MDBK cell culture with a note on enhancing effect of preincubation with chicken spleen cells
Jong-Yil Chai,Soon-Hyung Lee,Woong-Heum Kim,* and Chong Ku Yun*
Department of Parasitology and Institute of Endemic Diseases, College of Medicine,Seoul National University, Seoul 110-460, Korea.
Abstract
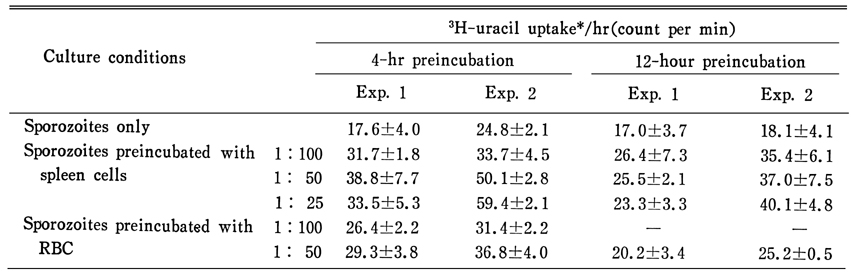
Eimeria tenella, an intracellular protozoan parasite infecting the epithelial cells of the ceca of chickens, causes severe diarrhea and bleeding that can lead its host to death. It is of interest that E. tenella first penetrate into the mucosal intraepithelial lymphocytes (IEL) before they parasitize crypt or villous epithelial cells. This in vitro study was undertaken to know whether the penetration of E. tenella into such a lymphoid cell is a beneficial step for the parasite survival and development. Three sequential experiments were performed. First, the in vitro established bovine kidney cell line, MDBK cells, were evaluated for use as host cells for E. tenella, through morphological observation. Second, the degree of parasite development and multiplication in MDBK cells was quantitatively assayed using radioisotope-labelled uracil (3H-uracil). Third, the E. tenella sporozoites viability was assayed after preincubation of them with chicken spleen cells. E. tenella oocysts obtained from the ceca of the infected chickens were used for the source of the sporozoites. Spleen cells (E) obtained from normal chickens (FP strain) were preincubated with the sporozoites (T) at the E:T ratio of 100:1, 50:1 or 25:1 for 4 or 12 hours, and then the mixture was inoculated into the MDBK cell monolayer. Morphologically the infected MDBK cells revealed active schizogonic cycle of E. tenella in 3-4 days, which was characterized by the appearance of trophozoites, and immature and mature schizonts containing merozoites. The 3H-uracil uptake by E. tenella increased gradually in the MDBK cells, which made a plateau after 48-60 hours, and decreased thereafter. The uptake amount of 3H-uracil depended not only upon the inoculum size of the sporozoites but also on the degree of time delay (preincubation; sporozoites only) from excystation to inoculation into MDBK cells. The 3H-uracil uptake became lower as the preincubation time was prolonged. In comparison, after preincubation of sporozoites with spleen cells for 4 or 12 hours, the 3H-uracil uptake was significantly increased compared with that of control group. From the results, it was inferred that, although the penetration of E. tenella sporozoites into the lymphoid cells such as IEL is not an essential step, it should be at least a beneficial one for the survival and development of sporozoites in the chicken intestine.
Figures
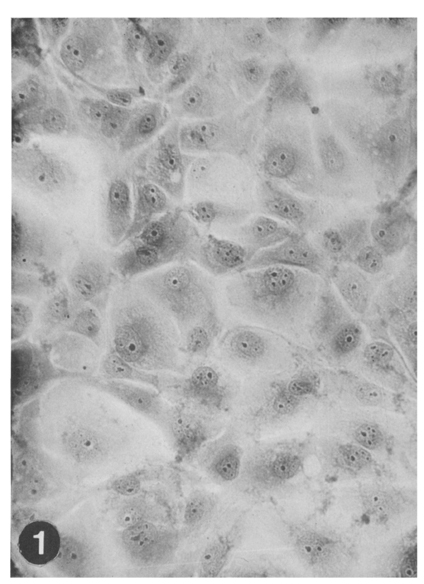
Fig. 1 MDBK (Madin Darby bovine kidney) cell monolayer, obtained 2 days after subculture with 1×105 cells/ml suspended in EBSS-EMEM medium, Hematoxylin-acid fuchsin-orange G (HT-AF-OG) stain, ×100.
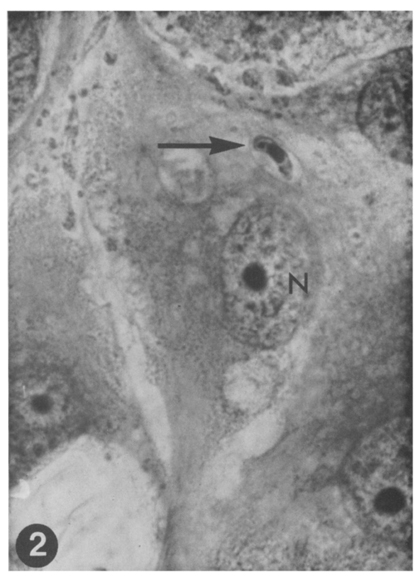
Fig. 2 A sporozoite of E. tenella (arrow) in the cytoplasm of MDBK cell surrounded by a parasitophorous vacuole of host origin, nearby the host cell nucleus(N). The sporozoite shows its nucleus at center and refractile vacuole at left terminal portion. One-day culture. HT-AF-OF stain, ×400.
Fig. 3 Ibid, another big sporozoite (arrow). The nucleus of sporozoite is seen in the center. One-day culture. HT-AF-OG stain, ×400.
Fig. 4 A round-up and an elongated form (trophozoite) of E. tenella in the cytoplasm of a MDBK cell, enveloped by surrounding parasitophorus vacuoles. One-day culture. HT-AF-OG stain, ×400.
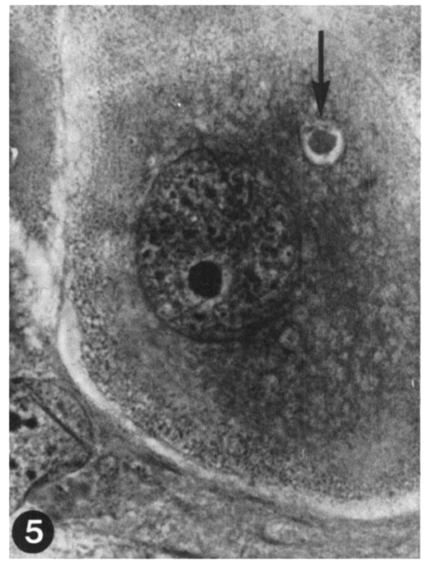
Fig. 5 A round-up trophozoite of E. tenella, which is probably dead or under degeneration. No nucleus nor globule is recognizable. Two-day culture. HT-AF-OG stain, ×400.
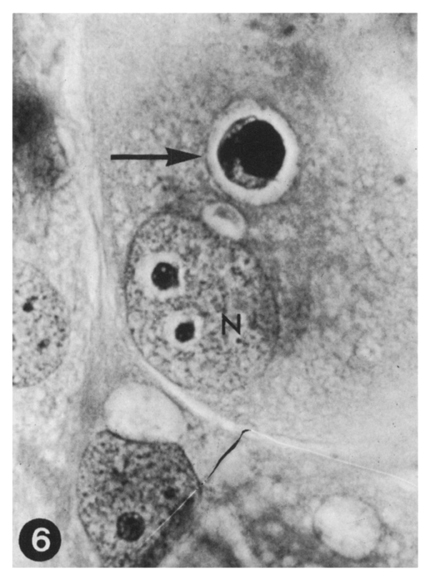
Fig. 6 A very big trophozoite of E. tenella, under further development. Host cell nucleus(N). Two-day culture. HT-AF-OG stain, ×400.
Fig. 7 An immature(developing) schizont of E. tenella in the cytoplasm of a MDBK cell. Its nuclear chromatin is divided into 7 or more. The parasite is occupying almost all of the host cell cytoplasm. Two-day culture. HT-AF-OG stain, ×400.
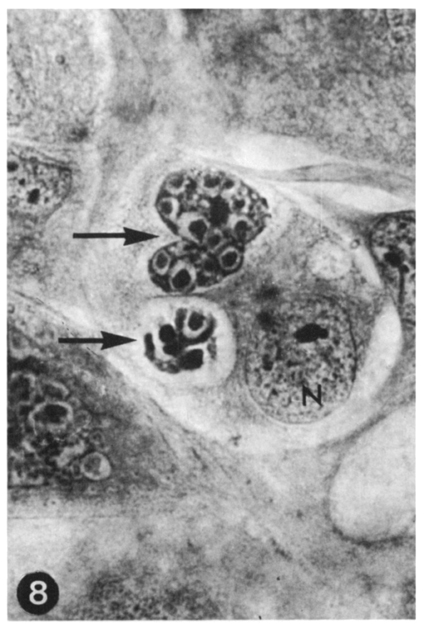
Fig. 8 Two immature schizonts of E. tenella (arrows), first generation. The upper one has distinctly been divided into about 15 merozoites. Two-day culture. HT-AF-OG stain, ×400.
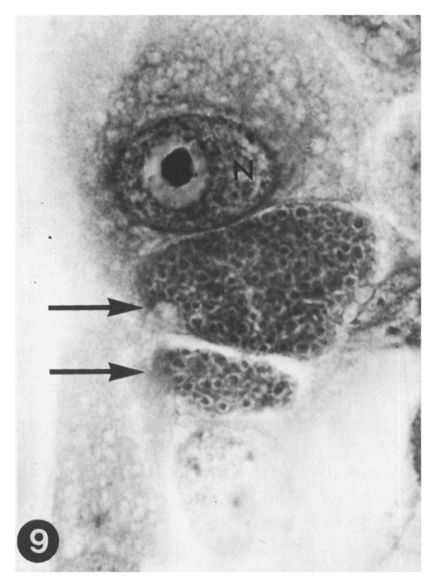
Fig. 9 Two immature schizonts of E. tenella (arrows), second generation. Numerous immature merozoites are seen at this stage. Host cell nucleus(N). Three-day culture. HT-AF-OG stain, ×400
Fig. 10 A mature schizoites (second generation) of E. tenella (arrows), containing numerous second generation merozoites. Three-day culture. HT-AF-OG stain, ×400.
Fig. 11 A fully mature schizont (large arrow) containing numerous mature merozoites (small arrows). The host cell is almost entirely occupied by the parasite and is about to rupture. Four-day culture. HT-AF-OG stain, ×400.
Fig. 12 Second generation merozoites (arrows) of E. tenella penetrating into new MDBK cells. Four-day culture. HT-AF-OG stain, ×400.
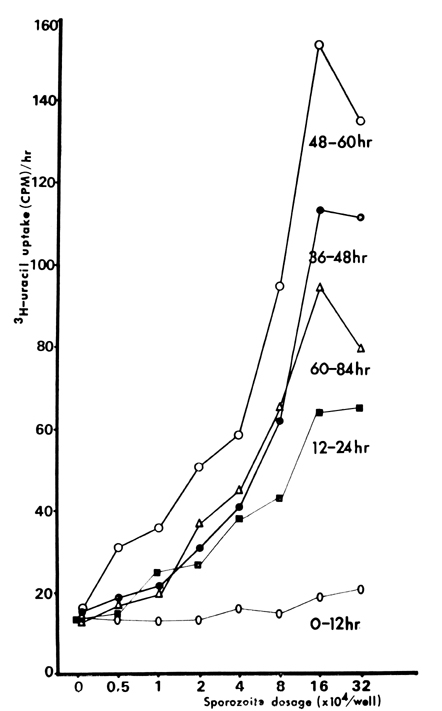
Fig. 13 3H-uracil uptake by E. tenella cultured in MDBK cell monolayer.
Fig. 14 3H-uracil uptake by E. tenella sporozoites preincubated in culture medium.
Tables
Table 1 3H-uracil uptake by E. tenella according to the dosage of sporozoite inoculum and time after inoculation into MDBK cell culture
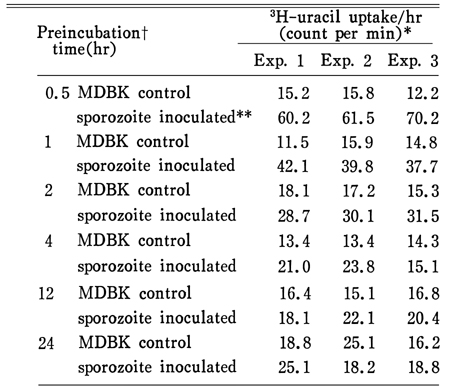
Table 2 3H-uracil uptake by E. tenella sporozoites according to preincubation time
Table 3 3H-uracil uptake by E. tenella preincubated for 4 or 12 hours
References
1.
Albright JW, Hatcher FM, Albright JF. Interaction between murine natural killer cells and trypanosomes of different species. Infect Immun 1984;44(2):315–319.
2.
Attallah AM, Lewis FA, Urritia-Shaw A, Folks T, Yeatman TJ. Natural killer cells (NK) and antibody-dependent cell-mediated cytotoxicity (ADCC) components of Schistosoma mansoni infection. Int Arch Allergy Appl Immunol 1980;63(3):351–354.
4.
Belosevic M, Faubert GM. Killing of Giardia muris trophozoites in vitro by spleen, mesenteric lymph node and peritoneal cells from susceptible and resistant mice. Immunology 1986;59(2):269–275.
5.
Chai JY, Lillehoj HS. Isolation and functional characterization of chicken intestinal intra-epithelial lymphocytes showing natural killer cell activity against tumour target cells. Immunology 1988;63(1):111–117.
8.
Crane MS, Schmatz DM, Stevens S, Habbersett MC, Murray PK. Eimeria tenella: in vitro development in irradiated bovine kidney cells. Parasitology 1984;88(Pt 3):521–530.
12.
Hatcher FM, Kuhn RE. Destruction of Trypanosoma cruzi by Natural killer cells. Science 1982;218(4569):295–296.
13.
Hauser WE Jr, Sharma SD, Remington JS. Natural killer cells induced by acute and chronic toxoplasma infection. Cell Immunol 1982;69(2):330–346.
14.
Hauser WE Jr, Sharma SD, Remington JS. Augmentation of NK cell activity by soluble and particulate fractions of Toxoplasma gondii. J Immunol 1983;131(1):458–463.
15.
Hauser WE Jr, Tsai V. Acute toxoplasma infection of mice induces spleen NK cells that are cytotoxic for T. gondii in vitro. J Immunol 1986;136(1):313–319.
16.
Herberman RB. Cell-mediated immunity to tumor cells. Adv Cancer Res 1974;19:207–263.
17.
Herberman RB. Natural killer (NK) cells and their possible roles in resistance against disease. Clin Immunol Rev 1981;1(1):1–65.
19.
Hunter KW, et al. Immunol Lett 1981;2:209–212.
20.
Kamiyama T, Hagiwara T. Augmented followed by suppressed levels of natural cell-mediated cytotoxicity in mice infected with Toxoplasma gondii. Infect Immun 1982;36(2):628–636.
21.
Kirkpatrick CE, Farrell JP. Splenic natural killer-cell activity in mice infected with Leishmania donovani. Cell Immunol 1984;85(1):201–214.
22.
Kogut MH, Lange C. Interferon-gamma-mediated inhibition of the development of Eimeria tenella in cultured cells. J Parasitol 1989;75(2):313–317.
23.
Lawn AM, Rose ME. Mucosal transport of Eimeria tenella in the cecum of the chicken. J Parasitol 1982;68(6):1117–1123.
24.
Lillehoj HS. Immune response during coccidiosis in SC and FP chickens. I. In vitro assessment of T cell proliferation response to stage-specific parasite antigens. Vet Immunol Immunopathol 1986;13(4):321–330.
25.
Lillehoj HS, Ruff MD. Comparison of disease susceptibility and subclass-specific antibody response in SC and FP chickens experimentally inoculated with Eimeria tenella, E. acervulina, or E. maxima. Avian Dis 1987;31(1):112–119.
26.
Merino F, Cruz I. Natural killer activity in experimental cutaneous leishmaniasis. Int Arch Allergy Appl Immunol 1984;73(4):347–351.
27.
Nauss KM, Pavlina TM, Kumar V, Newberne PM. Functional characteristics of lymphocytes isolated from the rat large intestine. Response to T-cell mitogens and natural killer cell activity. Gastroenterology 1984;86(3):468–475.
28.
Peirce MA. A simple staining technique to demonstrate the sporozoite refractile globule in coccidian parasites. Parasitology 1980;80(3):551–554.
29.
Pittilo RM, Ball SJ, Norton CC, Gregory MW. Penetration of chicken peripheral blood lymphocytes by Eimeria acervulina sporozoites. Ann Trop Med Parasitol 1986;80(5):569–571.
30.
Quan PC, Rager-Zisman B, Wittner M, Tanowitz HB. Interferon and natural killer cells in murine Chagas disease. J Parasitol 1983;69(6):1164–1166.
32.
Rosenberg EB, Herberman RB, Levine PH, Halterman RH, McCoy JL, Wunderlich JR. Lymphocyte cytotoxicity reactions to leukemia-associated antigens in identical twins. Int J Cancer 1972;9(3):648–658.
33.
Schmatz DM, Crane MS, Murray PK. Eimeria tenella: parasite-specific incorporation of 3H-uracil as a quantitative measure of intracellular development. J Protozool 1986;33(1):109–114.
34.
Skamene E, Stevenson MM, Lemieux S. Murine malaria: dissociation of natural killer (NK) cell activity and resistance to Plasmodium chabaudi. Parasite Immunol 1983;5(6):557–565.
35.
Solomon JB. Natural cytotoxicity for Plasmodium berghei in vitro by spleen cells from susceptible and resistant rats. Immunology 1986;59(2):277–281.
37.
Tagliabue A, Befus AD, Clark DA, Bienenstock J. Characteristics of natural killer cells in the murine intestinal epithelium and lamina propria. J Exp Med 1982;155(6):1785–1796.
38.
Wilson AD, Stokes CR, Bourne FJ. Morphology and functional characteristics of isolated porcine intraepithelial lymphocytes. Immunology 1986;59(1):109–113.