Generation and Immunity Testing of a Recombinant Adenovirus Expressing NcSRS2-NcGRA7 Fusion Protein of Bovine Neospora caninum
Article information
Abstract
Neospora caninum is the etiologic agent of bovine neosporosis, which affects the reproductive performance of cattle worldwide. The transmembrane protein, NcSRS2, and dense-granule protein, NcGRA7, were identified as protective antigens based on their ability to induce significant protective immune responses in murine neosporosis models. In the current study, NcSRS2 and NcGRA7 genes were spliced by overlap-extension PCR in a recombinant adenovirus termed Ad5-NcSRS2-NcGRA 7, expressing the NcSRS2-NcGRA7 gene, and the efficacy was evaluated in mice. The results showed that the titer of the recombinant adenovirus was 109TCID50/ml. Three weeks post-boost immunization (w.p.b.i.), the IgG antibody titer in sera was as high as 1:4,096. IFN-γ and IL-4 levels were significantly different from the control group (P<0.01). This research established a solid foundation for the development of a recombinant adenovirus vaccine against bovine N. caninum.
Neosporosis is an intracellular protozoan disease caused by Neospora caninum, which parasitizes dogs, cattle, sheep, and other mammals [1]. Dogs are the final host of N. caninum, and many mammals are intermediate hosts [4]. Neosporosis mainly causes miscarriage of dams, stillbirths, neonatal movement disorders, and neurologic diseases [2,3]. N. caninum is widespread in 30 countries, including the UK, USA, Australia, New Zealand, and South Korea. The infection rate is generally from 10 to 40%, and occasionally up to 80% [5-9]. There are reports involving N. caninum in Beijing, Qinghai, Xinjiang, Hebei, Jilin, and Heilongjiang in China [10-14]. With the frequent trading of cattle, endemic areas of neosporosis are gradually expanding. Neosporosis has been one of the important diseases affecting the development of the cattle industry [15,16].
Recently, some valuable surface proteins, such as NcSAG1 and NcSRS2, have been identified. NcSAG1 and NcSRS2 are mainly expressed in tachyzoites and bradyzoites using immunoblotting and immunofluorescence staining methods [17-19]. NcSRS2, a surface protein, plays an important role in the adhesion to and invasion of host cells, but anti-NcSRS2 mAbs can prevent this process [20,21]. The NcSRS2 gene has been expressed in baculovirus successfully, and can be used in the diagnosis and as a vaccine candidate gene [22,23]. Nishikawa et al. [22,23] used recombinant vaccinia viruses expressing the NcSRS2 and NcSAG1 genes to inoculate unmated BALB/c mice intraperitoneally. The recombinant vaccinia viruses induced mice to produce protective immunity, and the immune effect induced by NcSRS2 was better than that induced by NcSAG1 [24]. The NcGRA7 gene encodes the dense granule protein, which mainly regulates the parasitophorous vacuole (PV) to provide nutrition for the growth and proliferation of the parasite [25]. Jenkins et al. [26] investigated the immune effect of plasmid DNA encoding NcGRA6 and NcGRA7, and confirmed that NcGRA7 can be used as an immunogen in the prevention of neosporosis.
N. caninum tachyzoites of the Jilin strain were cultured in vero cells at 37℃ in an atmosphere containing 5% CO2 and purified as described previously [27]. N. caninum NcSRS2 and NcGRA7 genes were amplified using primers P1 and P2, and P3 and P4, respectively (Table 1). EcoRI and Xba I restriction enzyme sites were added to the 5´ ends of P1 and P4, respectively. The linker was added to the 5´ ends of P2 and P3, respectively. PCR amplification was performed in a 25 µl reaction system containing 1 µl of each primer (10 µM each), 2 µl of extracted DNA, 2.5 µl of 10× PCR buffer, 2 µl of dNTP (2.5 mM each), and 0.25 µl of TaKaRa (Dalian, China) Taq (5 U/µl). The reactions were performed in a thermal cycler (Eppendof, Hamburg, Germany) with the following program: 1 pre-denaturing cycle for 5 min at 94℃; 10 denaturing cycles for 30 sec at 94℃; annealing for 45 sec at 58℃; extension for 60 sec at 72℃; and extension for 7 min at 72℃. P1 and P4 were added to the reaction system, followed by 30 denaturing cycles as mentioned above. PCR products were analyzed by 1.0% agarose gel electrophoresis stained with ethidium bromide (0.5 mg/ml). The 1,054 bp NcSRS2 gene was amplified using the P1 and P2 primers, and the 364 bp NcGRA7 gene was amplified using the P3 and P4 primers. Using the purified NcSRS2 and NcGRA7 genes as templates, the 1,463 bp NcSRS2-NcGRA7 fusion gene was amplified using P1 and P4 primers (Fig. 1).

Primers for amplifying the NcSRS2 and NcGRA7 genes of N. caninum
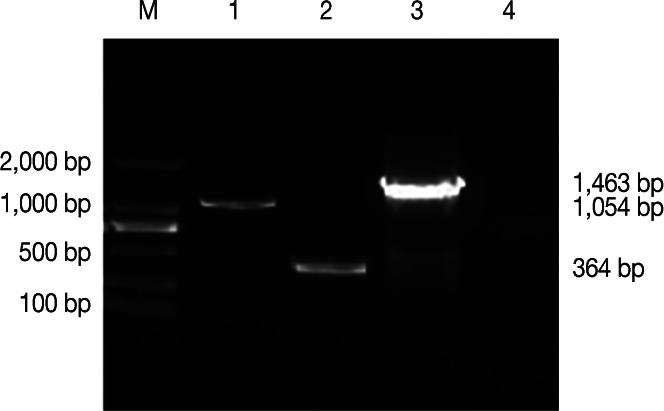
Results of PCR and SOE-PCR amplification. Lane M, DNA marker DL2,000; Lane 1, PCR products of the NcSRS2 gene; Lane 2, PCR products of the NcGRA7 gene; Lane 3, PCR products of the NcSRS2-NcGRA7 fusion gene; Lane 4, water control.
The NcSRS2-NcGRA7 fusion gene was purified and cloned into the pMD-18T simple vector, and referred to as pMD18-NcSRS2-NcGRA7. The recombinant plasmid was digested with EcoRI, Xba I, and subcloned into the adenovirus transfer vector, pCR259 (Qbiogene, Carlsbad, California, USA), digested with the same restriction enzymes, and termed pCR259-NcSRS2-NcGRA7. The recombinant adenovirus transfer vector was transformed into HighQ-1 Transpose-ADTM 294 competent cells (Qbiogene), a bacterial strain carrying the Transpose-ADTM 294 plasmid and a plasmid encoding a transacting Tn7 transposase. Through the Tn7-based transposition, the recombinant Transpose-Ad™ plasmid carrying the NcSRS2-NcGRA7 fusion gene was constructed. The recombinant Transpose-Ad™ plasmid was transformed into HighQ-1™ competent cells (Qbiogene) to be purified and amplified. The purified recombinant Transpose-Ad™ plasmid was linearized with Pac I and transfected into QBI-HEK 293 cells (Qbiogene,) using Lipofectamine™ 2000 (Invitrogen, Carlsbad, California, USA) to generate a recombinant adenovirus expressing the NcSRS2-NcGRA7 fusion gene product, and termed Ad5-NcSRS2-NcGRA7. The cytopathic effect (CPE) of infected QBI-HEK 293 cells was regularly observed. Seven days after the transfection, a CPE was observed in the cells. Twelve days after the transfection, the CPE was as high as 90% in the cells (Fig. 2).
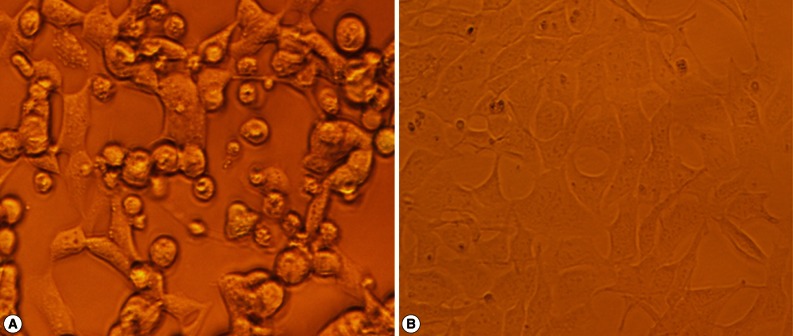
QBI-HEK 293 cells packaging Ad5-NcSRS2-NcGRA7. (A) QBI-HEK 293 cells packaging Ad5-NcSRS2-NcGRA7 (×1,000). (B) normal QBI-HEK 293 cells (×1,000).
QBI-HEK 293 cells, presenting >90% CPE, were collected. The genomic DNA was extracted. PCR amplification was carried out using primers P1 and P4. The 1,463 bp target band was amplified as expected (Fig. 3). QBI-HEK 293 cells, packaging recombinant adenovirus expressing the NcSRS2-NcGRA7 fusion gene product, were subcultured in 12-well plates, dried naturally, and fixed with cold acetone for 10 min. N. caninum-positive sera were added onto the fixed cells and incubated for 1 hr at 37℃ followed by thrice-washing with PBS. Then, fluorescein isothiocyanate (FITC)-labeled anti-bovine IgG was applied to the cells and incubated for 1 hr at 37℃, followed by thrice-washing with PBS. The cells were observed and photographed under a fluorescence microscope. Green fluorescence was observed in Ad5-NcSRS2-NcGRA7-infected 293 cells under the fluorescence microscope (Fig. 4A), but not in empty plasmid-infected 293 cells (Fig. 4B).
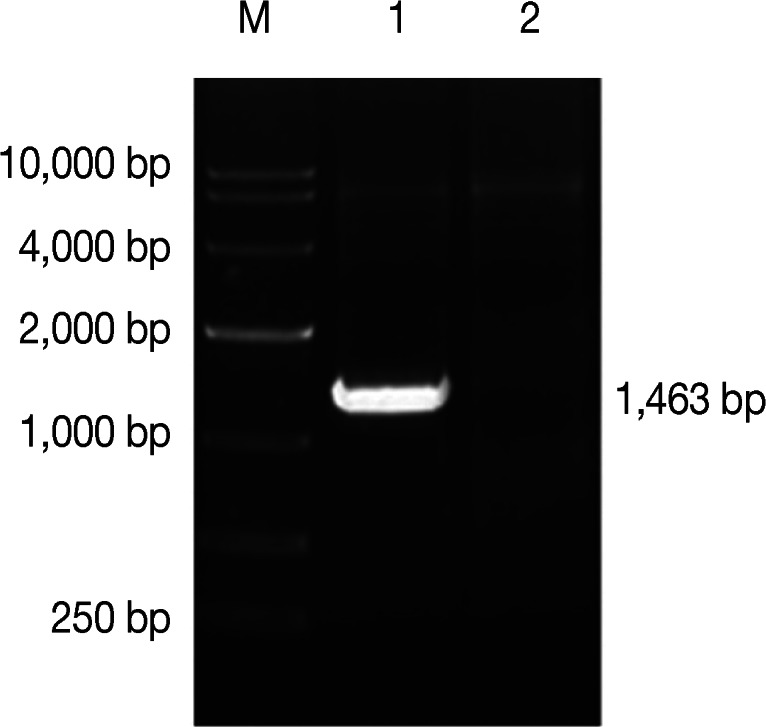
PCR identification of Ad5-NcSRS2-NcGRA7. Lane M, DNA marker is DL10,000; Lane 1, PCR products of Ad5-NcSRS2-NcGRA7; Lane 2, empty plasmid control.
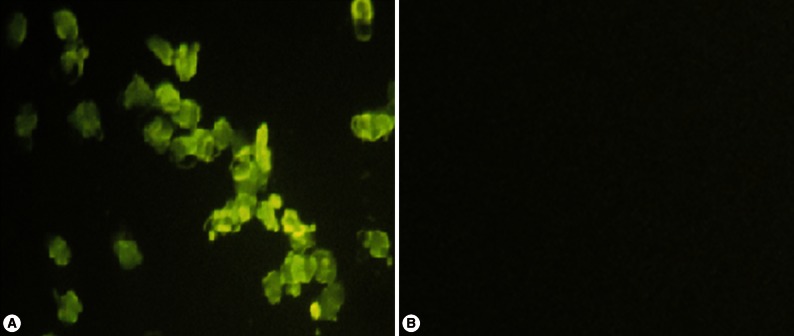
IFA detection of the expression of NcSRS2-NcGRA7 fusion gene. (A) Ad5-NcSRS2-NcGRA7-infected 293 cells (×400). (B) empty plasmid-infected 293 cells (×400).
QBI-HEK 293 cells, packaging recombinant adenovirus expressing the NcSRS2-NcGRA7 fusion gene product, were disrupted by repeated freezing and thawing, and ultrasonic methods. Cell lysates were separated by SDS-PAGE and transferred to nitrocellulose (NC) membranes by Western blotting. The transferred membranes were then placed in blocking solution (3% skim milk in PBS) at 4℃ overnight. The transferred membranes were incubated with N. caninum-positive sera and anti-bovine IgG conjugated with horseradish peroxidase at 37℃ for 2 hr followed by thrice-washing with PBS. AEC (3-Amino-9-ethylcarbazole) was added for color development. The predicted size of the NcSRS2-NcGRA7 fusion protein, which was about 51 kDa, was observed in Ad5-NcSRS2-NcGRA7-infected 293 cells, but not in empty plasmid-infected 293 cells (Fig. 5).
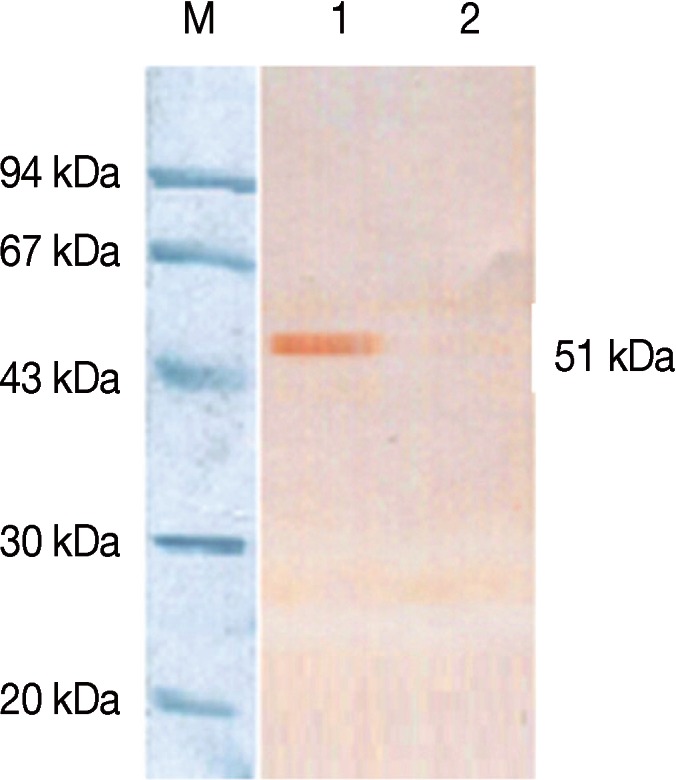
Western blotting analysis of the expression of the NcSRS2-NcGRA7 fusion gene. Lane M, Low molecular weight marker; Lane 1, expression products of Ad5-NcSRS2-NcGRA7-infected 293 cells; Lane 2, empty plasmid control.
QBI-HEK 293 cells were seeded onto 96-well cell plates for 48 hr. The cell nutrient solution was discarded, and the diluted virus solution (10-fold serial dilution with Dulbecco's minimum essential medium containing 2% sera) was inoculated into the cells. Each dilution was inoculated into 4 wells (100 µl diluted virus solution/well). The plate was incubated in a CO2 incubator at 37℃. The CPE of infected QBI-HEK 293 cells was regularly noted, and the number of CPE wells in each dilution was counted. By calculating, the titer of the recombinant adenovirus, Ad5-NcSRS2-NcGRA7, was 109 TCID50/ml.
Vaccines, using live viruses as vectors, can induce an immune response against the target antigen and will be a promising new vaccine [28]. The recombinant replication-defective adenovirus has a high efficiency of gene transfer and can induce protective immunity, including humoral and cellular immunity [29].
To evaluate the efficacy of the recombinant adenovirus which we had constructed, 6-week-old healthy BALB/c mice (n=30) were randomly divided into 3 groups. Intramuscular vaccinations in lower extremities were performed. BALB/c mice in group 1 were inoculated twice with 109 TCID50 Ad5-NcSRS2-NcGRA7 at 3-week intervals. The group 2 BALB/c mice were immunized twice with 109 TCID50 Ad5-GFP at 3-week intervals. The group 3 BALB/c mice were immunized twice with 50 µl of PBS. Sera were collected 7, 14, and 21 days after the first immunization. An indirect ELISA was applied to detect the IgG antibody titer in sera from week 1 post-primary immunization (PPI). At week 3 PPI, the IgG titers were as high as 1:2,048, and at week 3 post-boost immunization (PBI), the titers were 1:4,096, while specific IgG was not detected in Ad5-GFP and PBS groups (Fig. 6). Cytokine ELISA kits were used to detect the levels of IFN-γ and IL-4, in accordance with the manufacturer's instructions. Statistical analyses were performed using PASW Statistics (version 18).
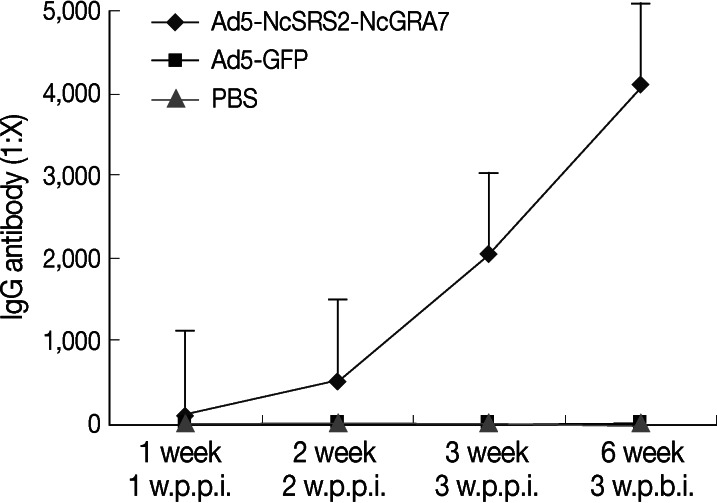
IgG antibody titer induced by Ad5-NcSRS2-NcGRA7 in mice as determined by indirect ELISA. BALB/c mice were inoculated at 3-week intervals 3 times, and sera were collected at week 2 after each immunization for antibody titer determination. The IgG antibody titers in sera were measured by indirect ELISA. Data were analyzed by 1-way ANOVA. w.p.p.i., week post-prime immunization; w.p.b.i., week post-boost immunization.
The levels of IgG, IFN-γ, and IL-4 were significantly elevated (P<0.05) from week 1 PPI to week 3 PBI. The levels of IFN-γ and IL-4 in the Ad5-NcSRS2-NcGRA7 group were significantly higher (P<0.01) than in Ad5-GFP and PBS groups (Fig. 7A, B). Thus, Ad5-NcSRS2-NcGRA7 could induce BALB/c mice to produce significant cellular immune responses.
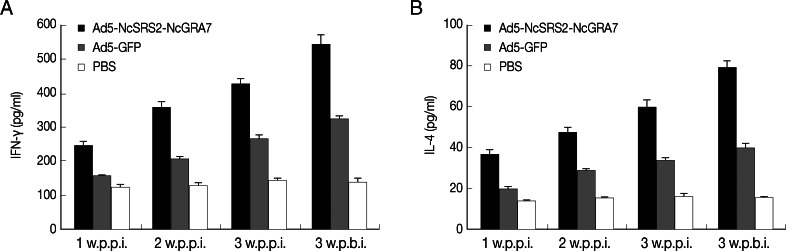
Levels of IFN-γ (A) and IL-4 (B) in sera of immunized BALB/c mice. BALB/c mice were inoculated and the levels of IFN-γ (A) and IL-4 (B) were measured using IFN-γ and IL-4 ELISA kits. Data were analyzed by 1-way ANOVA. w.p.p.i., week post-prime immunization; w.p.b.i., week post-boost immunization.
Antigenic proteins, which play an important role in the life cycle of N. caninum, should be taken into accounts in the design of vaccines. These include the surface protein, micro-line protein, and dense granule protein [17,25,30]. The NcSRS2 gene, which has an open reading frame (ORF) of 1,206 bp, encodes 401 amino acids and expresses a protein of 43 kDa, mediates parasite invasion into host cells [20]. The NcGRA7 gene, encoding a dense granule protein and expresses a protein of 33 kDa, provides nutrition for parasites and promotes their proliferation [25]. It was reported that NcSRS2 and NcGRA7 proteins can be used for diagnosis and vaccines [22,26,31].
At present, expression of the parasite exogenous gene by viral vectors has been widely applied in the parasitology field at home and abroad, and recombinant vaccines using virus as vectors have achieved remarkable results [32]. Adenoviruses are double-stranded DNA viruses and can infect a wide variety of cells. They have the highest replication efficiency among viral vectors and deliver their genome to the nucleus with extremely high efficiencies. Adenoviruses are currently the most promising vectors [33]. The Transpose-Ad™ Adenovirus Vector System is a Tn7-based transposon system and can complete homologous recombination of the target gene in Escherichia coli.
In the current study, we constructed recombinant adenovirus Ad5-NcSRS2-NcGRA7 based on the Transpose-Ad™ system. In homologous recombination, the recombinant adenovirus transfer plasmid pCR259-NcSRS2-NcGRA7 should complete the recombination and purification in HighQ-1 Transpose-AD™294 and HighQ-1™ competent cells. Moreover, the key technology to construct recombinant adenovirus Ad5-NcSRS2-NcGRA7 by the Transpose-Ad™ system is that homologous recombination-positive clones must be filtered on LB plates which contain Cm, Amp, and Tc at the same time.
The representative cytokines secreted by Th1 and Th2 cells are IFN-γ and IL-4, respectively. The main function of IFN-γ is to inhibit the infection of pathogens and their invasion by activating the immune system [34]. B lymphocytes, activated by IL-4, proliferate, differentiate, and secrete specific antibodies [35]. During N. caninum infection, Th1 and Th2 immune responses are involved in the host resistance to the parasite [36]. When the body was stimulated by exogenous antigens, T-cell subsets would be activated to secrete a variety of cytokines. Th1 cells secrete IFN-γ, IL-2, and TNF-α. These cytokines can significantly inhibit the propagation of N. caninum [37]. Th2 cells secrete IL-4, IL-5, and IL-13, and IL-4 can significantly inhibit the vertical transmission of N. caninum [38]. The secretions of IFN-γ and IL-4 in vivo are mutually restrictive in immune responses [39]. The levels of IFN-γ and IL-4 were both enhanced and thus could be balanced by Ad5-NcSRS2-NcGRA7 in immunized animals. This indicated that Ad5-NcSRS2-NcGRA7 was able to induce cellular immune responses of immunized mice.
In our study, we spliced NcSRS2 and NcGRA7 genes using overlap-extension PCR method. Hereafter, we constructed a recombinant adenovirus termed Ad5-NcSRS2-NcGRA7, expressing the NcSRS2-NcGRA7 gene. The recombinant adenovirus was immunogenic to mice.
ACKNOWLEDGMENTS
We are grateful to Xuenan Xuan of the National Research Center for Protozoan Diseases, Obihiro University of Agriculture and Veterinary Medicine in Japan for fruitful discussion and access to materials. The project was supported by the National Natural Science Foundation (31160501), China.