Activation of MAPK Is Required for ROS Generation and Exocytosis in HMC-1 Cells Induced by Trichomonas vaginalis-Derived Secretory Products
Article information
Abstract
Trichomonas vaginalis is a flagellated protozoan parasite that causes vaginitis and cervicitis in women and asymptomatic urethritis and prostatitis in men. Mast cells have been reported to be predominant in vaginal smears and vaginal walls of patients infected with T. vaginalis. Mitogen-activated protein kinase (MAPK), activated by various stimuli, have been shown to regulate the transcriptional activity of various cytokine genes in mast cells. In this study, we investigated whether MAPK is involved in ROS generation and exocytotic degranulation in HMC-1 cells induced by T. vaginalis-derived secretory products (TvSP). We found that TvSP induces the activation of MAPK and NADPH oxidase in HMC-1 cells. Stimulation with TvSP induced phosphorylation of MAPK and p47phox in HMC-1 cells. Stimulation with TvSP also induced up-regulation of CD63, a marker for exocytosis, along the surfaces of human mast cells. Pretreatment with MAPK inhibitors strongly inhibited TvSP-induced ROS generation and exocytotic degranulation. Finally, our results suggest that TvSP induces intracellular ROS generation and exocytotic degranulation in HMC-1 via MAPK signaling.
INTRODUCTION
Trichomonas vaginalis is a flagellated protozoan parasite that causes vaginitis and cervicitis in women and asymptomatic urethritis and prostatitis in men [1-4]. Trichomoniasis is the most common curable sexually-transmitted infection [5]. More than 180 million people around the world are annually infected by this parasite [1,6]. In pregnant women, trichomonads are implicated in the premature rupture of membranes, premature delivery, and the delivery of low-birth-weight infants [7,8]. The pathogenesis induced by T. vaginalis infection begins with the attachment of the parasite to epithelial cells and subsequent stimulation of host cells by T. vaginalis-derived secretory products (TvSP), which induce pathogenic changes in host tissues, eliciting cytotoxicity and immune responses [9,10]. The trichomonad adhesion to vaginal epithelial cells and the release of proteolytic enzymes contained in TvSP are considered to be major factors responsible for disruption of vaginal epithelial layers [11,12].
Mast cells have been reported to be predominant in vaginal smears and vaginal walls of patients infected with T. vaginalis [13,14]. Mast cells can participate in direct killing of organisms by phagocytosis and reactive oxygen species (ROS) production [15]. ROS generation by membrane-localized NOX is a main cell signaling mechanism associated with tissue inflammation [16], and this may be dependent on the level of ROS production [17,18]. NOX2 is controlled by phosphorylation of cytosolic component p47phox [19]. Degranulation of mast cells is essential for host defense against parasitic infections, where they play an important role in innate and adaptive immune responses [20]. CD63 is a component of granular membranes and is used as a marker for exocytosis [16]. Recent studies showed that CD63 is known to be expressed in intracellular membranes, such as secretory lysosomes, including serotonincontaining granules [21,22].
The mitogen-activated protein kinase (MAPK) cascade is an important signaling pathway in immune responses [17,23,24]. The activation of MAPK cascades involved various biological responses of proliferation [25], migration [26], apoptosis [27,28], cytokine expression [29], and degranulation [23] in various cells. In mammalian cells, MAPK signaling comprises 3 major subfamilies, such as extracellular signal-regulated kinase (ERK), p38 MAP kinase, and c-Jun N-terminal kinase (JNK) [29-31]. Although previous studies have demonstrated that MAPK pathways are involved in the up-regulation of IL-8 production in neutrophils activated by T. vaginalis [29] and phosphorylation of p38 MAPK and ERK in T. vaginalis-infected HeLa cells [25], none have reported on the relationships between MAPK and TvSP-induced human mast cell activation. Therefore, in this study, we aimed to investigate the role of MAPK signaling on TvSP-induced ROS generation and exocytotic degranulation in HMC-1 cells.
MATERIALS AND METHODS
Reagents
The following reagents were purchased from Calbiochem (Gibbstown, New Jersey, USA): Platelet-activating factor (PAF) was purchased from Biomol (Plymouth Meeting, Pennsylvania, USA). Antibodies (Ab) specific for phospho-ERK1/2, phospho-p38MAPK, phospho-JNK1/2, total-JNK1/2, total-ERK1/2, total-p38 MAPK and β-actin were purchased from Cell Signaling Technology (Danvers, MA, USA). All other reagents were purchased from Sigma Chemical Company (St. Louis, Missouri, USA). Anti-p47phox antibody was purchased from BD Biosciences (San Jose, California, USA) and anti-phospho-p47phox antibody was kindly donated by Dr. Jamel El-Benna. Phycoerythrin (PE)-conjugated anti-human CD63 monoclonal antibody (mAb) was purchased from Biolegend (San Diego, California, USA). All other reagents were purchased from Sigma Chemical Company unless otherwise stated.
Cultivation of T. vaginalis trophozoites and preparation of secretory products (TvSP)
T. vaginalis was axenically subcultivated at 37˚C with Diamond’s trypticase yeast extract-maltose (TYM) medium with 10% heat inactivated horse serum (Gibco/Invitrogen, Gaithersburg, Maryland, USA) and 0.5% penicillin/streptomycin (Gibco/Invitrogen). T. vaginalis for the preparation of TvSP was obtained from logarithmically growing cells. To obtain various doses of TvSP for HMC-1 stimulation, trichomonads (1×107 or 2×105) were washed once with Hank’s balanced salt solution (HBSS) (Gibco/Invitrogen), resuspended in 1 ml HBSS, and incubated for 1 hr at 37˚C in order to collect TvSP. After incubation, culture supernatants were centrifuged for 10 min at 14,000 rpm and filtered through filter with 0.22 µm pores, yielding the TvSP used in this study. Protein concentrations were measured by the BCA protein assay using bovine serum albumin as a standard.
Cultivation of the human mast cell line
HMC-1 cells were used as a source of human mast cells. HMC-1 cells were axenically subcultivated at 37˚C with IMDM (Iscove’s medium) (Gibco/Invitrogen), containing 10% heat inactivated fetal bovine serum (FBS) (Biomedia, Foster City, California, USA) and 1% penicillin/streptomycin in a 5% CO2 incubator.
Stimulation of human mast cells and pretreatment with MAPK inhibitors
HMC-1 cells (1×105 or 5×105/well) seeded in 48- or 96-well tissue culture plates were stimulated for different time points with or without TvSP or PAF. In addition, HMC-1 cells (1×105/well) were pretreated for 30 min with DMSO (0.5%) at 37˚C as a control. Cells were also pretreated with inhibitor of ERK1/2 (PD98059), p38MAPK (SB203580), and JNK (SP600125) at 50 µM, before stimulation with TvSP or PAF.
Immunoblot analysis
HMC-1 cells (5×105/well) stimulated for different time points with or without TvSP or PAF in 24-well tissue culture plates in a CO2 incubator. After incubating for the indicated times, the reaction was stopped by brief centrifugation. The cells were lysed with lysis buffer (20 mM Tris-HCl, pH 7.5, 60 mM β-glycerophosphate, 10 mM EDTA, 10 mM MgCl2, 10 mM NaF, 2 mM DTT, 1 mM Na3VO4, 1 mM amidino-PMSF, 1% Nonidet P-40, and 5 μg/ml leupeptin) on ice for 30 min. After centrifuging at 12,000 g for 5 min, the supernatants were diluted in SDS-PAGE loading buffer and heated at 100˚C for 5 min. The samples were stored at -20˚C until ready for use. Samples were subjected to 8 or 10% SDS-PAGE followed by electrotransfer onto Immobilon P polyvinylidene fluoride membranes (Millipore, Billerica, Massachusetts, USA). The membranes were blocked with 5% nonfat dry milk in TBST at room temperature for 1 hr and then incubated with primary antibodies against phospho-SAPK/JNK (Thr183/Tyr185) antibody, phospho-p38 MAPK (Thr180/Tyr182) antibody, phospho- ERK1/2 p44/42 (Thr202/Tyr204) antibody, phospho-p47phox, SAPK/JNK antibody, p38 MAPK antibody, ERK1/2 antibody, p47phox antibody or β-actin antibody at 4˚C overnight. The membranes were subsequently soaked in HRP-conjugated anti-rabbit or goat IgG at room temperature for 1 hr. Immunoreactivity was detected using Lumi GLO (Cell Signaling).
Measurement of intracellular ROS generation in HMC-1 cells
Intracellular ROS accumulation in HMC-1 cells was measured using a green fluorescent probe, DCF-DA. HMC-1 cells (1×105/well) were pre-stained at 37˚C for 10 min with 1 μM of DCF-DA, which is rapidly oxidized to highly fluorescent DCF in the presence of intracellular H2O2. Cells were then incubated for up to 60 min with or without TvSP or PAF in 24-well tissue culture plates in a CO2 incubator. Cells were washed twice with washing buffer, and DCF fluorescence was measured by flow cytometry using a FACS Calibur TM (BD Biosciences). At least 10,000 gated events were analyzed for each sample.
Flow cytometric measurement of CD63 in HMC-1 cells
Exocytotic degranulation induced by TvSP was measured as the mean fluorescent intensity of surface CD63 expression levels by staining HMC-1 cells with PE-conjugated anti-human CD63 mAb. To obtain surface CD63 expression levels, HMC-1 cells (1×105) were stimulated with or without TvSP and PAF for up to 60 min. Cells were then incubated for 30 min at 4˚C with PE-conjugated anti-CD63 mAb or isotype control Ab, and washed 3 times with FACS buffer and then analyzed by flow cytometry. An appropriate irrelevant isotype control was used to measure nonspecific binding. The mean fluorescence intensity of CD63 expression for at least 10,000 cells from each sample was measured by flow cytometry.
Statistical analysis
The results are expressed as means±SD of 3 independent experiments. Student’s t-test was used for all statistical analyses, and all P-values <0.05 were considered statistically significant.
RESULTS
Effect of TvSP on MAPK activation in HMC-1
To determine whether TvSP could induce activation of MAPK (ERK1/2, p38 MAPK, and JNK) in human mast cells. As shown Fig. 1A, HMC-1 cells were stimulated with TvSP for various times 5, 15, and 30 min caused transient ERK1/2 activation. The phosphorylation of ERK1/2 induced by TvSP showed a peak at 5 min and declined after 15 min. As shown Fig. 1B, C, TvSP significantly increased the phosphorylation of p38 MAPK, and JNK in HMC-1 cells, compared to cells treated with medium alone. TvSP induces activation of p38 MAPK, and JNK in HMC-1 in a time-dependent manner. Platelet activating factor (PAF), as a positive control, also induced phosphorylation of ERK1/2, p38 MAPK, and JNK in HMC-1 cells. We found that TvSP-induced phosphorylation of ERK1/2 is transient compared with phosphorylation of p38 MAPK and JNK.
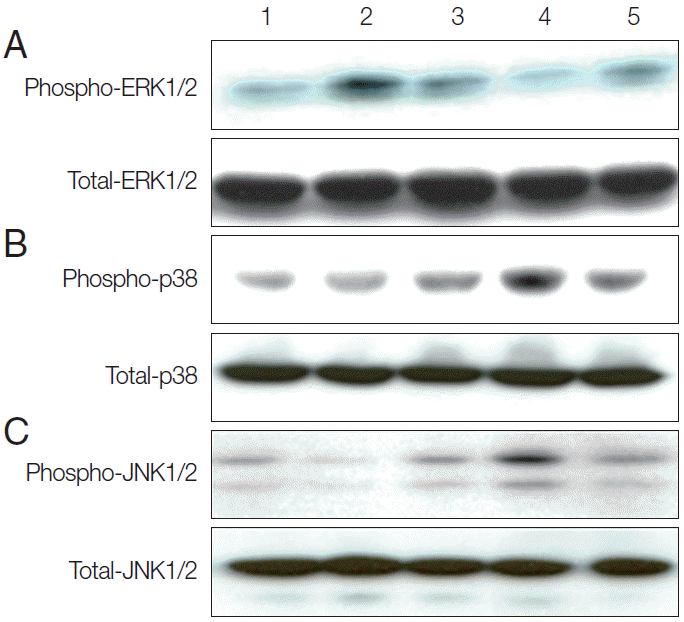
TvSP induces activation of ERK1/2 (A), p38 (B), and JNK (C) in HMC-1 cells. Stimulation with TvSP induces phosphorylation of MAPK in HMC-1 cells. HMC-1 cells (5×105/well) were incubated for 5, 15, and 30 min at 37°C with or without T. vaginalis (2×105/well) in transwell system for western blotting, with antibodies against phospho-ERK1/2, total ERK1/2; phospho-p38, total p38; phospho-JNK1/2, total-JNK1/2; (10% SDS-PAGE gel). Lane 1: HMC-1+medium, 30 min; Lane 2: HMC-1+TvSP, 5 min; Lane 3: HMC-1+TvSP, 15 min; Lane 4: HMC-1+TvSP, 30 min; Lane 5: HMC-1+PAF 1 μM, 30 min. PAF was treated as a positive control.
Effect of TvSP on NOX2 activation in HMC-1
Next, we attempted to examine whether TvSP can induce phosphorylation of p47phox in HMC-1 cells. Phosphorylation of p47phox is and important step for NOX2 activation [19]. As shown in Fig. 2, stimulation with TvSP strongly induced phosphorylation of p47phox after 15 min of stimulation, compared with medium alone. TvSP induces NOX activation in a time-dependent manner.
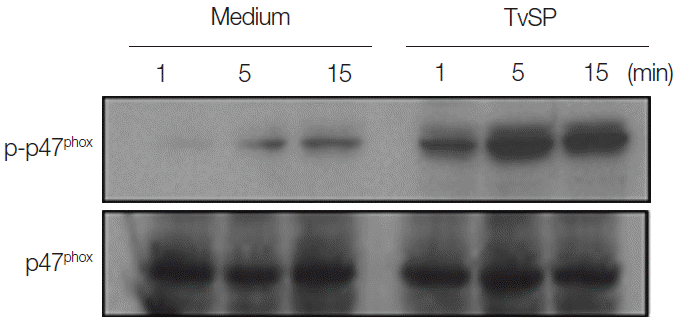
TvSP induces phosphorylation of p47phox in HMC-1 cells. HMC-1 cells were stimulated for 1, 5, and 15 min with or without TvSP and then blotted with antibodies against phospho-p47phox and total p47phox. The figure is representative of 3 experiments showing similar results.
Effect of TvSP on the intracellular ROS generation and exocytotic degranulation in HMC-1
We then performed flow cytometry analysis to assess whether TvSP induces intracellular ROS generation and exocytotic degranulation in HMC-1 cells. TvSP induced markedly up-regulation in intracellular ROS production (Fig. 3A) and exocytotic degranulation (Fig. 3B), compared with medium alone.
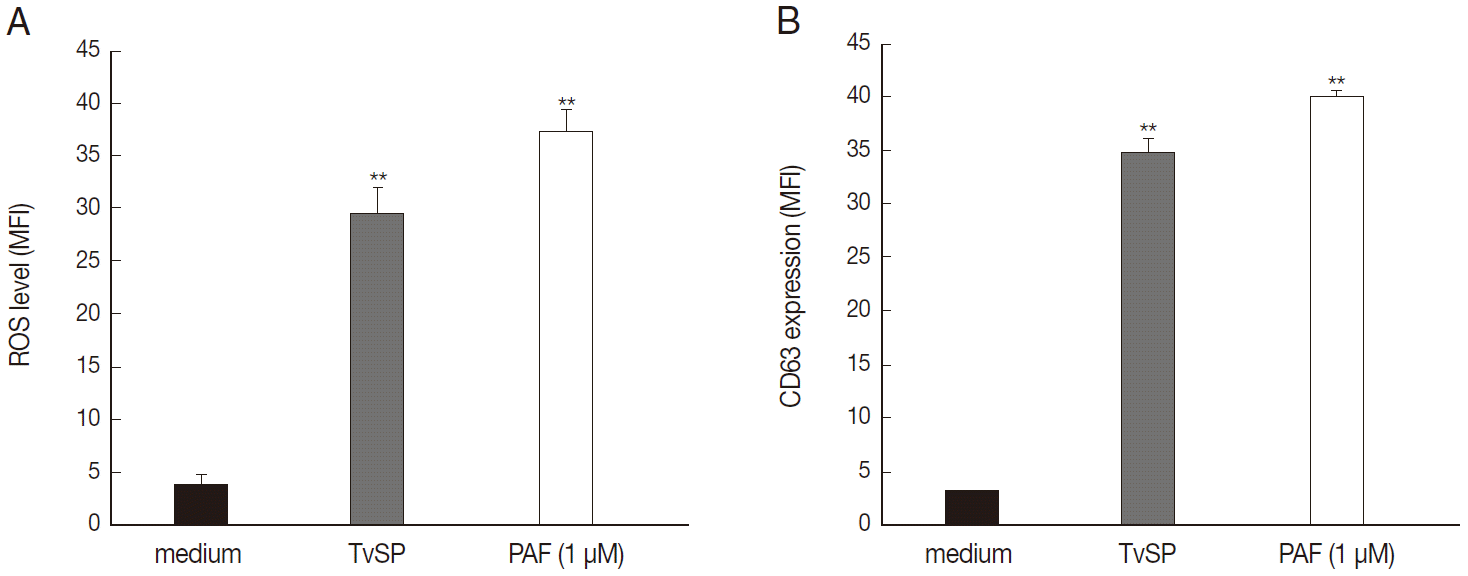
TvSP induces ROS generation and exocytotic degranulation in HMC-1 cells. HMC-1 cells (1×105) were stimulated for 1 hr with or without TvSP or PAF (1 μM). (A) Cells were stained with DCF-DA (1 μM) for 10 min. (B) CD63 expression. Data were analyzed by FACS and expressed the mean±SD from 3 independent experiments. **P<0.01 compared to control.
Role of MAPK on intracellular ROS generation and exocytotic degranulation in TvSP-stimulated HMC-1 cells
To determine whether MAPK activation is involved in TvSP-induced intracellular ROS generation and exocytotic degranulation in HMC-1 cells, we examined the inhibitory effects of 3 MAPK inhibitors, ERK1/2 (PD98059), p38MAPK (SB203580), and JNK (SP600125), in human mast cells. Pretreatment with inhibitor of ERK1/2 (PD98059), p38MAPK (SB203580), and JNK (SP600125) markedly inhibited ROS generation and CD63 expression in HMC-1 cells induced by TvSP. TvSP induced ROS generation and surface upregulation of the exocytosis marker protein CD63 in HMC-1 cells via MAPK-dependent signaling (Fig. 4).
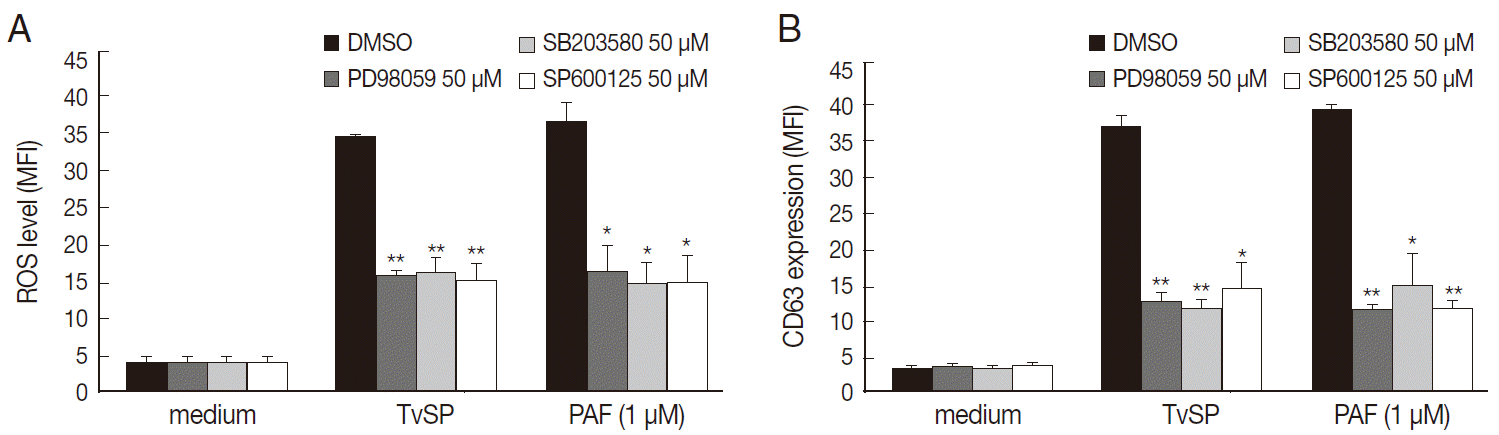
TvSP induces ROS generation and exocytosis in HMC-1 cells via activation of MAPK. HMC-1 cells (1×105) were pretreated with inhibitors of ERK1/2, p38 MAPK, and JNK (50μM) for 30 min before stimulation with TvSP. (A) Effect of inhibitors of MAPK on TvSP-induced intracellular ROS generation. (B) Effect of inhibitors of MAPK on TvSP-induced exocytotic degranulation in HMC-1 cells. Data were analyzed by FACS and expressed as the mean±SD from 3 independent experiments. Cytotoxicity of TvSP at the concentration used was observed. *P<0.05, **P<0.01 compared to the control.
DISCUSSION
In this study, we demonstrated for the first time the role of MAPK signaling in TvSP-induced ROS generation and exocytotic degranulation in HMC-1 cells. MAPK signaling pathway is activated in response to various external stresses including parasite and bacterial infections, hyperosmolarity, and UV irradiation [32]. However, the role of MAPK on TvSP-induced human mast cell activation has not been reported.
Our results suggested that mast cells may play an important role in inflammatory reactions caused by T. vaginalis via MAPK signaling pathway. The amounts of phosphorylated p38 MAPK, and JNK increased at 5, 15, and 30 min, respectively. In particular, TvSP-induced phosphorylation of ERK1/2 is transient compared with phosphorylation of p38 MAPK and JNK. Previous studies showed that persistent activation of ERK1/2 is associated with cell death [23,28,31]. In our study, TvSP-induced transient ERK1/2 phosphorylation seems to be associated with cell activation.
NADPH oxidase (also called NOX) has been established as a key enzyme for generating intracellular ROS [19]. Recent studies have shown that phosphorylation of cytosolic subunit p47phox plays an essential role in NOX2 activity, which is required for ROS generation [16,19]. Therefore, we examined whether TvSP could induce phosphorylation of the NOX2 cytosolic component p47phox. Herein, phosphorylation of p47phox was increased by TvSP in a time dependent manner in HMC-1 cells, in accordance with a previous study which showed that NADPH oxidase activity and excessive ROS generation could be involved in inflammation and destruction, through increased p47phox-Ser345 phosphorylation [26]. Additionally, NADPH oxidase-derived intracellular ROS generation has been shown to mediate T. vaginalis-induced apoptosis of human neutrophils [31], migration of neutrophils [29] and monocytes [4], and activation of transcription factors in HMC-1 cells [3], leading to an inflammatory response in the prostate epithelium [4].
Thus, we attempted to determine whether TvSP induces intracellular ROS production and exocytotic degranulation in HMC-1 cells by flow cytometry analysis. TvSP induced markedly increases in intracellular ROS production and exocytotic degranulation in HMC-1 cells, compared with medium alone. Previous studies evidenced that T. vaginalis can induce ROS generation in human neutrophils [31], migration of neutrophils [29], monocytes [4], and activation of transcription factors in HMC-1 cells [3], lead to an inflammatory response in prostate epithelium [4]. Taken together, our results suggest that TvSP-induced intracellular ROS generation and degranulation is associated with tissue inflammation.
Mast cells contain potent mediators, including histamine, serotonin, heparin, proteases, leukotrienes, and cytokines, and they are preformed and stored in granules from which they are released upon mast cell activation in a process called degranulation or regulated exocytosis [15,21,33]. Degranulation of mast cells has been reported in allergic inflammation and during infections with parasites. For example, the absence of CD63 results in a significant decrease in degranulation of mast cells [21], which translates into a reduction of acute allergic reactions in vivo, thus identifying CD63 as an important component of allergic inflammations [22]. A previous study also demonstrated that mast cells stimulated with TvSP release histamine and TNF-α [15]. In our study, we showed that mast cells play a role in the inflammation caused by T. vaginalis via exocytotic degranulation, as TvSP significantly increased exocytotic degranulation in HMC-1 cells. Nevertheless, we were unable to discern which signaling mechanism is involved in TvSP-induced human mast cell activation during inflammatory responses after acute T. vaginalis infection.
Previous studies have demonstrated that intracellular ROS generated from phorbol myristate acetate (PMA)-stimulated HMC-1 cell contribute to the production of inflammatory cytokines via p38 MAPK [30]. Additionally, phosphorylation of p47phox has been shown to play a key role in regulation of TNF-α-induced ROS generation via MAPK signaling in endothelial cells [34]. MAPK is also known to mediate p47phox phosphorylation on Ser345 in TNF-α-treated neutrophils [26]. Our study showed that MAPK activation plays a major role in ROS generation in HMC-1 cells induced by TvSP. In addition, we showed effects of 3 kinds of MAPK inhibitors such as PD98059, SB203580, and SP600125 on TvSP-induced exocytotic degranulation in HMC-1 cells. Pretreatment of HMC-1 cells with inhibitors of MAPK markedly suppressed CD63 expression in HMC-1 cells induced by TvSP. This result suggests that MAPK-dependent CD63 expression plays a key role in the exocytotic degranulation of HMC-1 cells induced by TvSP.
Previous studies have demonstrated that T. vaginalis-secreted lipid mediator LTB4 induces IL-8 production in mast cells via BLT-dependent activation of NF-kB and CREB [3]. Further, NOX2-derived ROS regulates surface trafficking of BLT1 during exocytotic degranulation in human eosinophils stimulated with LTB4 [16]. Accordingly, we plan to investigate whether lipid mediator LTB4, a component of TvSP, induces host BLT-dependent activation of MAPK in human mast cells.
In conclusion, we showed that MAPK signaing is important for TvSP-induced ROS generation and exocytotic degranualtion in human mast cells. These results support our understanding of the pathogenesis and host-parasite relationships in T. vaginalis infections.
Acknowledgements
This work was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education, Science and Technology (2011-0021634).
Notes
We have no conflict of interest related to this work.