Mass Death of Predatory Carp, Chanodichthys erythropterus, Induced by Plerocercoid Larvae of Ligula intestinalis (Cestoda: Diphyllobothriidae)
Article information
Abstract
We describe here the mass death of predatory carp, Chanodichthys erythropterus, in Korea induced by plerocercoid larvae of Ligula intestinalis as a result of host manipulation. The carcasses of fish with ligulid larvae were first found in the river-edge areas of Chilgok-bo in Nakdong-gang (River), Korea at early February 2016. This ecological phenomena also occurred in the adjacent areas of 3 dams of Nakdong-gang, i.e., Gangjeong-bo, Dalseong-bo, and Hapcheon-Changnyeong-bo. Total 1,173 fish carcasses were collected from the 4 regions. To examine the cause of death, we captured 10 wondering carp in the river-edge areas of Hapcheon-Changnyeong-bo with a landing net. They were 24.0-28.5 cm in length and 147-257 g in weight, and had 2-11 plerocercoid larvae in the abdominal cavity. Their digestive organs were slender and empty, and reproductive organs were not observed at all. The plerocercoid larvae occupied almost all spaces of the abdominal cavity under the air bladders. The proportion of larvae per fish was 14.6-32.1% of body weight. The larvae were ivory-white, 21.5-63.0 cm long, and 6.0-13.8 g in weight. We suggest that the preference for the river-edge in infected fish during winter is a modified behavioral response by host manipulation of the tapeworm larvae. The life cycle of this tapeworm seems to be successfully continued as the infected fish can be easily eaten by avian definitive hosts.
Ligula intestinalis (Cestoda: Diphyllobothriidae) is one of the tapeworm species inhabiting the intestine of fish-eating birds. This tapeworm is widely distributed in the world, and has a complicate 3-host life cycle like Diphyllobothrium and Spirometra spp. [1]. Unembryonated eggs discharged in the feces of avian hosts develop and hatch out in the water as the coracidium. The ciliated coracidia freely swimming in the water are eaten by the first intermediate host, i.e., crustacean copepods, and then develop into procercoids in the abdominal cavity of copepods within 2-3 weeks. Copepodes with procercoids are eaten by the second intermediate hosts, zooplankton-eating fish. Infected procercoids develop into plerocercoids in the abdominal cavity of the fish hosts. The plerocercoid larvae stay more than 10 months in the fish host, whereas in the definitive host, fish-eating birds, the larvae quickly mature into adults having gravid proglottids which stay no longer than 5-6 days [1,2].
There have been many studies on the infection status of ligulid larvae in the second intermediate fish hosts [2-8]. However in the Republic of Korea (Korea), only 1 study was performed. Moreover, it was a brief manuscript submitted to the 38th Korea Science Exhibition by Ryu and Lee [9]. After that, a local news paper reported that more than 80% of pale chubs, Zacco platypus, in Hoengseong-dam were highly infected with these tapeworm larvae in December 2003. On the other hand, it has been known that the plerocercoid larvae of L. intestinalis induce endocrine disruption, in particular, parasitic castration [10-14], growth suppression, morphological modification [15,16], and mortality increase in the fish [17,18] as well as modified behavioral responses [4,19-22]. Recently, we observed a mass death of predatory carp, Chanodichthys erythropterus, infected with the plerocercoid larvae of L. intestinalis in the 4 sites of Nakdong-gang (‘gang’ means river). Thus, in the present study, we intended to describe this exceptional ecological phenomenon and the cause of death of predatory carp from the aspects of host-parasite relationship.
Carcasses of predatory carp with ligulid larvae were first found in the river-edge areas of Nakdong-gang located below 1 km site of Chilgok-bo (‘bo’ means dam) in Chilgok-gun, Gyeongsangbuk-do, Korea at 7 February 2016 (Fig. 1). After then, this ecological phenomena also occurred in the adjacent areas of 3 dams of Nakdong-gang, i.e., Gangjeong-bo, Dalseong-bo, and Hapcheon-Changnyeong-bo, as well as Chilgok-bo. A total of 1,173 fish carcasses were collected from the 4 regions during about 40 days. The fatal status of predatory carp infected with the L. intestinalis plerocercoids was designated in Table 1.

The predatory carp, Chanodichthys erythropterus, found in the river-edge areas of Chilgok-bo in Nakdong-gang (River), Korea. (A) A slowly swimming carp with an extruding larva. (B) An exhausted one just before dying with an escaping larva. (C) Four carcasses of fish with escaping holes of larvae. All scale bars are 10 cm.

To examine the cause of death, we captured 10 wondering carp in the river-edge area of Hapcheon-Changnyeong-bo with a landing net. They were 24.0-28.5 (av. 25.6) cm in length and 147-257 (av. 178) g in weight, and had 2-11 (5.3) plerocercoid larvae in the abdominal cavity. Their digestive organs were slender and empty without intestinal contents, and reproductive organs were not observed at all, while the plerocercoid larvae occupied almost all spaces of the abdominal cavity under the air bladders (Fig. 2). The proportion in weight of larvae per fish was 14.6-32.1% (av. 23.0%) of the total body weight. The larvae were ivory-white, long slender with the blunt anterior end (scolex part), 21.5-63.0 (av. 38.1) cm in length and 6.0-13.8 (av. 8.6) g in weight (Fig. 3).

The predatory carp captured in the river-edge areas of Hapcheon-Changnyeong-bo with a landing net. (A) A complete one with the bulging abdomen before necropsy. (B) Two fish opened their abdominal cavity to examine the cause of death. Plerocercoid larvae (white mass) of L. intestinalis are occupying almost all space of the abdominal cavity under the air bladders. The digestive organs of fish are slender and empty without intestinal contents (arrow marks). Fig.
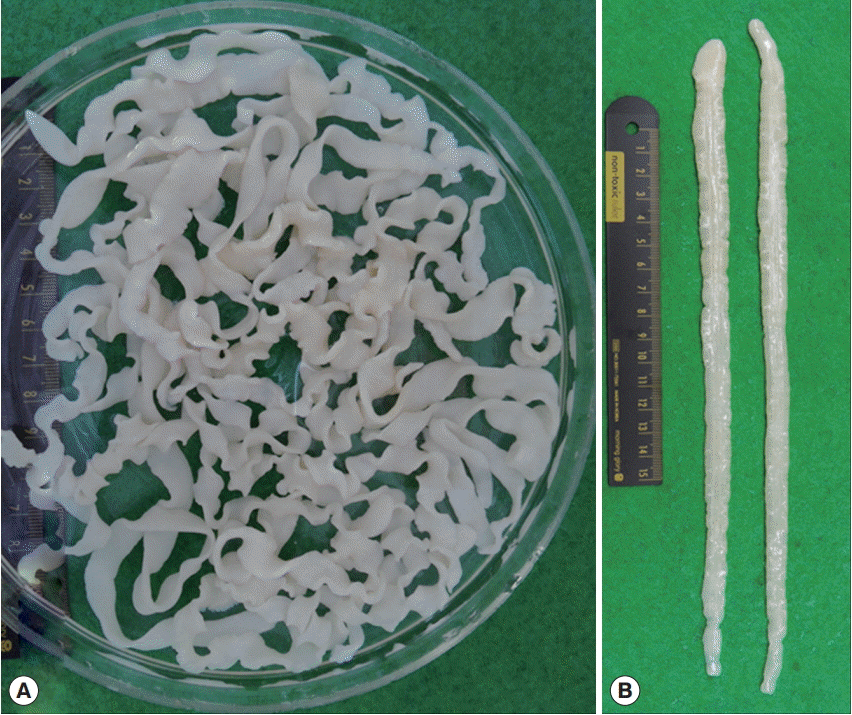
Plerocercoid larvae of L. intestinalis recovered in the abdominal cavity of fish. (A) All larvae recovered from a heavily infected fish. (B) Two larvae showing their gross morphology with the blunting anterior end (scolex part).
Host manipulation by parasites is known as one of the strategies for parasite survival. Various examples have been studied with aspects of host-parasite relationships [23]. As an interesting example, mice infected with Toxoplasma gondii are easily eaten by the definitive host, the cat, because they become lack of phobia response against the predator [24]. In case of L. intestinalis, it has been known that the plerocercoid larvae induce endocrine disruption that can lead to parasitic castration [10-14], growth suppression, morphological modification [15,16], mortality increase in the host fish [17,18], and modified behavioral responses [4,19-22]. Therefore, the preference for the river-edge in L. intestinalis-infected fish during winter season is suggested to be a modified behavioral response similar to host manipulation by ligulid larvae. The infected carp slowly swimming in the river-edge area can be easily eaten by avian definitive hosts (migratory birds) during their visiting season, winter, and the life cycle of this tapeworm will be successfully continued.
During the life cycle of this tapeworm, the plerocercoid stage is most conspicuous in aspects of host-parasite relationships. The long-lasting inhabitation and development of L. intestinalis in the abdominal cavity will give considerable effects on the fish host, i.e., physical damage, morphological modifications, growth retardation, hematological changes, immune response disturbance, endocrine disruption, and behavioral changes [4,19-27]. Among these effects, the physiological function related to reproduction was frequently studied using the fish-ligulid parasite model. The decrease of pituitary gonadotropins, sex steroids, and brain aromatase expressions, as well as the inhibition of gonad maturation and development were mainly revealed as the results of these studies [10-14,27]. In the present study, although we did not approach functionally, we were able to know the infertility of infected carp viewing from absence of their reproductive organs at necropsy of the fish, probably because of degeneration. On the other hand, it has been suggested that parasites can obviously influence host population in a density-dependent manner, if they reduce host survival and/or fecundity, and the abundance of potential hosts and the temporal dynamics of their occurrence strongly affect local host specificity [28]. Whether the mass death of predatory carp infected with plerocercoid larvae of L. intestinalis affects the host population or not? It is one of the problems to be solved in the near future from ecological aspects of fish and parasites.
As the second intermediate host of this tapeworm, cyprinid fish are typically reported from a variety of location worldwide. The roach, Rutilus rutiles, is the most well known fish host in European countries, and it has been most frequently studied as a suitable fish model. In African continent, Barbus species have been broadly examined to know the infection status of plerocercoid larvae of L. intestinalis [4,7]. In Asian countries, including China and Korea, the cultrinid fish (subfamily Cultrinae), in particular, Hemiculter bleekeri, was reported as the fish intermediate host of L. intestinalis [9]. By the present study, another species of cultrinid fish, C. erythropterus, has been confirmed as a suitable second intermediate host of this tapeworm in Korea. Accordingly, until now, total 4 species of freshwater fish, i.e., Zacco platypus, Carassius auratus, H. bleekeri, and C. erythropterus, are listed as the second intermediate hosts of L. intestinalis in Korea.
The carcasses of predatory carp found in the river-edge areas had the escape holes of larvae just behind the thoracic fin. The plerocercoid larvae probably escaped through these holes when fish hosts were dead. At necropsy of fish, all visceral organs, including the digestive and reproductive ones of fish, were severely atrophied, and the plerocercoid larvae occupied almost all spaces of the abdominal cavity under air bladders. The parasite weight reached about 23.0% of the total body weight of the fish. The proportion of larval weight per fish was somewhat higher than those of the previous studies. Loot et al. [2] reported that average 6 plerocercoid larvae were infected in 3-year-old roach, R. rutilus, from a gravel pit in River Garonne in France, and their weight was 17.5% weight of fish infected. Britton et al. [4] reported about 20% weight of ligulid larvae in Barbus lineomaculatus from Lake Baringo in Kenya.
The ligulid larval infections occurred more frequently in fish from lakes and dams rather than rivers. In case of Korea, they were detected in fish from 5 lakes in Chungcheongnam-do and Hoengseong-dam in Gangwon-do [9], and 4 dams in Nakdong-gang in this study. Most of the studies in other countries were performed with fish from lakes and dams except a few [4-8]. What kinds of ecological factors act in the life cycle of this tapeworm? The preference of stagnated water environment for the life cycle of this tapeworm should be investigated in the near future.
In conclusion, the plerocercoid larvae of L. intestinalis recently caused mass fatality of the fish host, C. erythropterus, in Korea. The host-parasite relationships between the fish and the tapeworm seem to be evolutionally well maintained, and the larvae were grown well in the abdominal cavity of fish without negative host responses. Ecological factors and the reason why the mass death of predatory carp occurred exceptionally in the dams of Nakdong-gang should be clarified in the near future. In addition, L. intestinalis is an avian parasite but not a human parasite, although Joyeux and Noyer [29] described it from the vomitus of a Romanian person, and thus some textbooks and websites treat this as a human tapeworm [30]. L. intestinalis should have strong host specificity for fish and birds during a long evolutional process.
Acknowledgements
We thank Jung-A Kim and Hee-Ju Kim, Department of Parasitology and Tropical Medicine, Gyeongsang National University School of Medicine, Jinju, Korea, for their help in fish examinations.
Notes
The authors declare that they have no conflict of interest related to this article.