Prevalence of Centrocestus formosanus Metacercariae in Ornamental Fish from Chiang Mai, Thailand, with Molecular Approach Using ITS2
Article information
Abstract
The prevalence of Centrocestus formosanus metacercariae was investigated in ornamental fish purchased from a pet shop in Chiang Mai, Thailand, including Carassius auratus (goldfish), Cyprinus carpio (Koi), Poecilia latipinna (Sailfin Molly), Danio rerio (Zebrafish), and Puntigrus tetrazona (Tiger barb). The parasite species was identified by the morphology of worms as well as by a molecular approach using ITS2. The results showed that 50 (33.3%) of 150 fish examined were infected with the metacercariae. The highest prevalence was found in C. auratus (83.3%), and the highest intensity was noted in C. carpio (70.8 metacercariae/fish). The most important morphological character was the presence of 32–34 circumoral spines on the oral sucker. The phylogenetic studies using the rRNA ITS2 region revealed that all the specimens of C. formosanus in this study were grouped together with C. formosanus in GenBank database. This is the first report on ornamental fish, C. carpio, P. latipinna, D. rerio, and P. tetrazona, taking the role of second intermediate hosts of C. formosanus in Thailand. Prevention and control of metacercarial infection in ornamental fish is urgently needed.
Centrocestus formosanus (Nishigori, 1924) (Heterophyidae) was described from Taiwan and now known to distribute widely in Asia [1]. The adult worm lives in the intestine of fish-eating birds and mammals [2–4]. Human cases infected with C. formosanus were reported in Lao PDR and Vietnam [4,5]. Pleurolophocercous cercariae are shed from the thiarid snail (Melanoides tuberculata) [1,6]. The metacercariae are found on the gill of several freshwater fish species, such as Puntius brevis, Hampala dispar, Puntius gonionotus, Puntius meiacanthus, Cyclocheilichthys armatus, Anabas testudineus, and Henicorhynchus siamensis [7–9]. In addition, the ornamental fish were reported to be infected with C. formosanus [2]. The parasite infection causes a problem in fish culture and leads to a reduction of fish production in aquaculture [1,10]. The presence of metacercariae on the gills of fish could be one of the reasons for the death of fish [2]. In Thailand, 2 species of Centrocestus, including C. formosanus and C. caninus, were reported; however, C. caninus is regarded as a synonym of C. formosanus [4].
The adult stage of this fluke was found in humans from Chiang Mai and Chiang Rai Provinces, northern Thailand [11,12]. The metacercariae were found in several species of freshwater fish [7,13–17]. However, there were few studies on metacercarial infection in ornamental fish species. Thus, the objective of this study was to determine the prevalence and species identification of Centrocestus in ornamental fish purchased from a pet shop in Chiang Mai Province, northern Thailand through morphological and molecular studies.
Total 150 ornamental fish, including 30 Carassius auratus, 30 Cyprinus carpio, 30 Poecilia latipinna, 30 Danio rerio, and 30 Puntigrus tetrazona, were collected from a pet shop in the Mueang District, Chiang Mai Province, northern Thailand during May–June 2016. The metacercariae were investigated on the gill of fish under a stereomicroscope, and then all the mature metacercariae (with X-shape excretory bladder) were collected for further studies. Some of the metacercariae were fed to chicks (1-day old), and after 7 days the adult stages were collected from their small intestines. Some other metacercariae were used for molecular studies. The adult stage was used to make a permanent slide according to Boonchot et al. [18]. All specimens were fixed with 4% formalin, stained with hematoxylin, dehydrated in alcohol series, and finally mounted in permount.
C. formosanus genomes were extracted by Chelex (Fluka, Sigma-Aldrich, St. Louis, Missouri, USA) following Caron et al. [19]. DNA products were amplified with ITS2 region. The reactions were performed in a Thermal Cycler machine (Little Genius, Bioer Technology, Tokyo, Japan). The primer combination; forward 3S (5′-GGT ACC GGT GGA TCA CTC GGC TCG TG-3′) and reverse BD2 (5′-TAT GCT TAA ATT CAG CGG GT-3′) were performed in PCR for ITS2 gene. The PCR conditions were as follows: 2 min initial denaturation at 94°C, followed by 35 cycles of 1 min DNA denaturation at 94°C, 1 min primer annealing at 60°C, 1 min extension at 72°C, and 7 min for final extension at 72°C. PCR products were tested by gel electrophoresis with DNA Dye NonTox (Applichem, Darmstadt, Germany) stain. After gel electrophoresis, PCR products were purified and sent to sequence analysis. The sequence was analyzed by ClustalW in MEGA software version 6.0 [20]. The sequence of C. formosanus was compared and checked by BLAST program on National Center for Biotechnology Information database for species confirmation and gathering of essential sequences for phylogenetic analysis. ITS2 sequences of C. formosanus, Centrocestus sp., Haplorchis taichui, Haplorcoides sp., Stellantchasmus falcatus, Paramphistomum epiclitum, Fasciola gigantica, and Heterakis gallinarum from GenBank were aligned with C. formosanus from this study. The phylogenetic tree was constructed using the MEGA software version 6.0. The data was analyzed by character analysis (maximum-likelihood) and distance analysis (neighbor-joining) with 1,000 bootstrap values.
Total 50 (33.3%) of 150 ornamental fish examined were infected with metacercariae of C. formosanus. The highest prevalence was found in C. auratus (83.3%), and the highest intensity of infection was in C. carpio (av. 70.8 metacercariae per fish). The results of other fish are shown in Table 1. The measurements of adult worms are shown in Table 2. The morphology of C. formosanus from 5 ornamental fish was almost similar. The metacercariae were oval shaped, with X-shaped excretory bladder and 32–34 circumoral spines surrounding the oral sucker. The adult flukes were pyriform shaped, and the tegument was covered with scale like spines. All adult worms originating from C. auratus possessed exclusively 34 circumoral spines (Fig. 1A1, A2), whereas those worms originating from other fish were armed with 32 spines arranged in 2 rows (Fig. 1B1, B2). They were both regarded as C. formosanus.
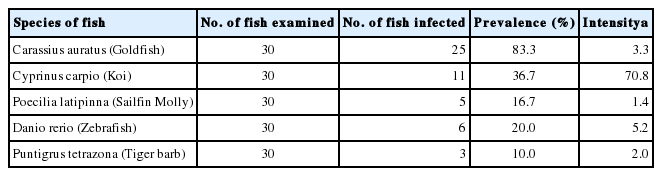
Prevalence and intensity of infection with Centrocestus formosanus metacercariae in ornamental fish species from Chiang Mai, Thailand
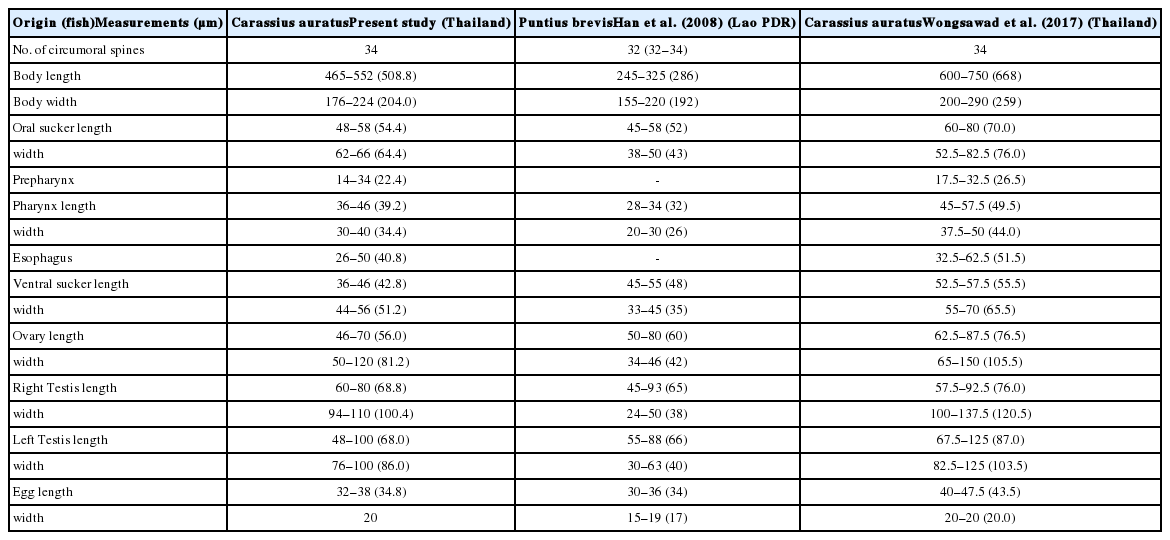
Measurements (range and mean) of Centrocestus formosanus specimens originating from Carassius auratus (n=10) compared with 2 previous studies
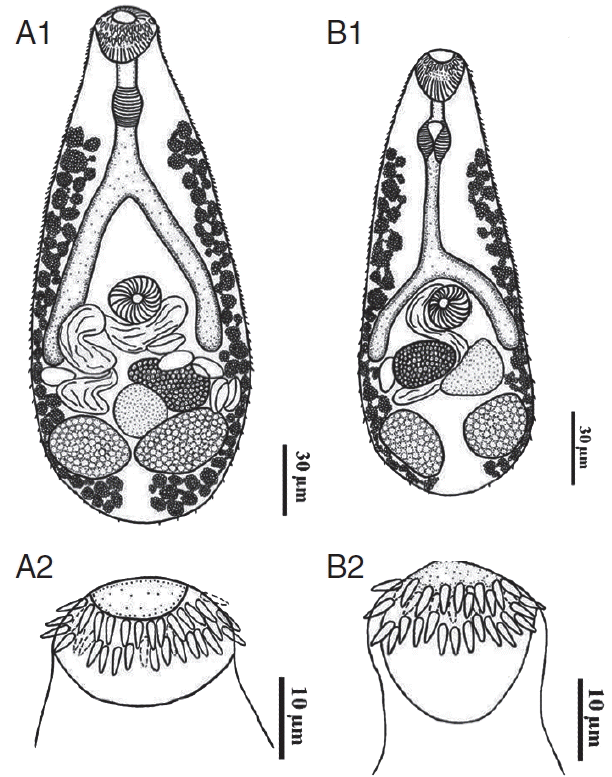
Line drawing of an adult Centrocestus formosanus originating from C. auratus (A1) showing 34 circumoral spines around the oral sucker (A2) and another originating from C. carpio (B1) showing 34 circumoral spines around the oral sucker (B2).
The BLAST results of C. formosanus originating from C. auratus of this study showed 99% similarity with 5.8S rRNA gene (partial sequence), ITS2 of C. formosanus GenBank accession no. KJ630863. The phylogenetic trees were reconstructed based on a maximum-likelihood and neighbor-joining methods with bootstrap values of 1,000 replicates. The results from both methods showed that the topology is similar (Fig. 2) to that of C. formosanus originating from C. auratus [25] which was grouped with Centrocestus sp. and C. formosanus from GenBank with 100% and 98% bootstrap values, respectively.
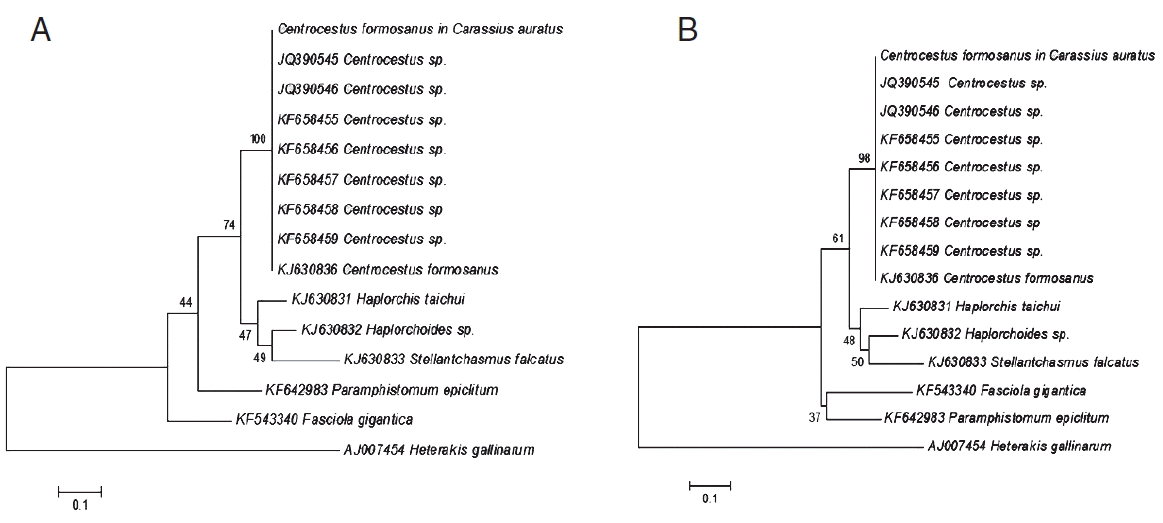
Phylogenetic trees of C. formosanus originating from C. auratus in Chiang Mai, Thailand. (A) A phylogenetic tree analyzed by maximum-likelihood (ML) method using the MEGA program software version 6.0 with 1,000 bootstrap values. (B) Another tree analyzed by neighbor-joining (NJ) method using the MEGA program software version 6.0 with 1,000 bootstrap values.
C. formosanus metacercariae were found in ornamental fish from many countries, such as Mexico, Australia, Denmark, and Iran [21–24]. In Thailand, the metacercariae of C. formosanus have been reported in several fish species, such as Macrognathus siamensis, P. gonionotus, P. brevis, Thynnichthys thynnoides, Puntioplites proctozysron, Esomus metallicus, A. testudineus, Parambassis siamensis, and Hampala macrolepidota [14,25,26]. However, this fluke is not well known in ornamental fish in Thailand, and in Chiang Mai Province, it has been reported only in C. auratus [25,27]. In this study, the metacercariae of C. formosanus were found in 5 species of ornamental fish. The results elucidated that ornamental fish can serve as the second intermediate host for C. formosanus. In addition, C. formosanus metacercariae were found for the first time in C. carpio, D. rerio, P. latipinna and P. tetrazona in Thailand.
The species confirmation of C. formosanus was based on morphological and molecular methods. The unique character of C. formosanus is the number of circumoral spines around the oral sucker. They commonly have 32 circumoral spines arranged in 2 rows around the oral sucker [1,4,8,18,24,28], whereas Centrocestus armatus has 42–44 circumoral spines [3,29,30]. In the present study, there were 32 circumoral spines in worms originating from 4 fish species (C. carpio, P. latipinna, D. rerio, and P. tetrazona) which agreed to previous studies [1,2,24,28]. The worms originating from C. auratus fish had exclusively 34 circumoral spines which also resembled previous studies [8,21]. Thus, our specimens of 2 different origins were both considered morphologically to be C. formosanus. Molecular studies of our specimens using ITS2 by maximum-likelihood and neighbor-joining methods revealed a high relationship with C. formosanus from GenBank. They were separated from other heterophyid flukes and also from P. epiclitum, F. gigantica, and H. gallinarum (out group). The results of morphological and molecular studies were accorded.
This is the first report of C. formosanus metacercariae in C. carpio, P. latipinna, D. rerio, and P. tetrazona ornamental fish. The phylogenetic trees showed high relationships of our specimens with C. formosanus from GenBank database. Infection of these fish with C. formosanus metacercariae should be prevented and controlled. It can also help to reduce the motility rate of fish before export or sell to customers.
ACKNOWLEDGMENTS
We would like to thank Environmental Science Research Center (ESRC), Chiang Mai University, Chiang Mai, Thailand. Special thanks to the staff of the Applied Parasitology Research Laboratory, Department of Biology, Faculty of Science, and Chiang Mai University for the facility supported. We are also thankful to the Science and Technology Research Institute, Chiang Mai University for financial support.
Notes
CONFLICT OF INTEREST
We have no conflict of interest related to this study.