Protective efficacy of vaccination with Neospora caninum multiple recombinant antigens against experimental Neospora caninum infection
Article information
Abstract
Protective efficacy of vaccination with Neospora caninum multiple recombinant antigens against N. caninum infection was evaluated in vitro and in vivo. Two major immunodominant surface antigens (NcSAG1 and NcSRS2) and two dense granule proteins (NcDG1 and NcDG2) of N. caninum tachyzoites were expressed in E. coli, respectively. An in vitro neutralization assay using polyclonal antisera raised against each recombinant antigen showed inhibitory effects on the invasion of N. caninum tachyzoites into host cells. Separate groups of gerbils were immunized with the purified recombinant proteins singly or in combinations and animals were then challenged with N.caninum. Following these experimental challenges, the protective efficacy of each vaccination was determined by assessing animal survival rate. All experimental groups showed protective effects of different degrees against experimental infection. The highest protection efficacy was observed for combined vaccination with NcSRS2 and NcDG1. Our results indicate that combined vaccination with the N. caninum recombinant antigens, NcSRS2 and NcDG1, induces the highest protective effect against N. caninum infection in vitro and in vivo.
INTRODUCTION
Neospora caninum is an obligatory intracellular protozoan parasite that infects a wide range of animals such as intermediate or definitive hosts, and has been recognized as an important causative agent responsible for significant economic and reproductive losses worldwide in the livestock industry, especially among dairy cattle (Dubey and Lindsay, 1996; Dubey, 2003). The main route of fetus infection by N. caninum is considered to involve the reactivation of latent tissue cysts or from the ingestion of oocysts during gestation. Currently, there is no effective method of controlling neosporosis except intensive farm management to reduce the likelihood of infection. Therefore, the development of a reliable vaccine against N. caninum infection is urgently needed to prevent infection and disease spread.
Although vaccination with whole lysates of tachyzoites was found to elicit effective protective immunity against the congenital transmission of N. caninum in experimental animals (Liddell et al., 1999; Andrianarivo et al., 1999; Choromanski and Block, 2000; Romero et al., 2004), the use of whole lysates of tachyzoites as commercial vaccines may not feasible, as they are expensive and time-consuming to produce and also contain many antigens that could elicit non-protective, immunosuppressive, or undesired immune responses. In this respect, extensive works have been undertaken to identify N. caninum specific antigens that elicit effective protective immunity and to develop reliable vaccines from them. These include parasite surface proteins and proteins of secretory organelles such as micronemes, rhoptries, and dense granules (Innes et al., 2002).
Two major surface proteins, NcSAG1 and NcSRS2, and two dense granule proteins, NcDG1 and NcDG2, of the parasite have been partially studied as potential vaccine candidates that are capable of inducing protective immunity against N. caninum infection, and specific antibodies against these proteins were found to inhibit the invasion of the tachyzoites into host cells in vitro (Howe et al., 1998; Augustine et al., 1999; Nishikawa et al., 2001; Cannas et al., 2003). However, it remains unclear as to which vaccine formulation is optimal for protection, since it is becoming apparent that different antigens are the targets of different immunological responses. Considering the complexities of the biology of N. caninum and of the host immune responses elicited by the parasite, an optimal vaccine for neosporosis could be expected to be a combination of the multiple antigens of N. caninum. The aim of this study was to determine if a combination of N. caninum specific antigens, namely, NcSAG1, NcSRS2, NcDG1, and NcDG2, can improve the protective efficacy against experimental N. caninum infection, and to further evaluate their applicabilities as vaccine candidates.
MATERIALS AND METHODS
Parasite
Tachyzoites of Neospora caninum (Kr2 strain) were propagated in Vero cells maintained in Eagle's minimum essential medium supplemented with 10% fetal bovine serum, 2 mM glutamine, and 20 µg/ml gentamicin (GIBCO-BRL, Grand Island, NY, USA). Parasites were harvested from their feeder cell cultures as described previously (Hempill and Gottstein, 1996). For purification of tachyzoites, the parasites and host cell debris were washed twice in cold phosphate-buffered saline (PBS, pH 7.2). The pellet obtained was resuspended in 2 ml of cold PBS and passed through a PD-10 column filled with Sephadex G25 M (Amersham Biosciences, Uppsala, Sweden) that had previously been equilibrated with PBS. The eluted, purified parasites were collected by centrifugation at 1,500 × g at 4℃ and resuspended in cold PBS. Purified tachyzoites were checked for viability using Trypan blue staining (0.25% in PBS). Whole lysates of tachyzoites were prepared as described previously (Andrianarivo et al., 1999)
Cloning, expression, and purification of the recombinant proteins
Total RNA was isolated from N. caninum tachyzoites using Trizol reagent (GIBCO-BRL). mRNA was purified by using Oligotex® mRNA purification kit (Qiagen, Valencia, CA, USA) by following the manufacturer's instructions. Single-stranded cDNA was synthesized from isolated mRNA using a cDNA synthesis kit (Boehringer Mannheim GmbH, Mannheim, Germany) and oligo-dT primer. To amplify each gene, PCR was performed using total cDNA and primer sets for each of NcSAG1, NcSRS2, NcDG1, and NcDG2 (Table 1). Amplification reactions were performed using the following conditions; 94℃ for 10 min, 35 amplification cycles (94℃ for 1 min, 55℃ for 1 min, and 72℃ for 2 min), followed by a 72℃ extension for 10 min. Amplified PCR products were analyzed on 1.2% agarose gel, purified from gels with a QIAquick gel extraction kit (Qiagen), and ligated into pCR2.1 vector (Invitrogen, Carlsbad, CA, USA), respectively. Each plasmid was transformed into competent E. coli TOP10 cells (Invitrogen) and the cloned genes were sequenced. Plasmid DNA containing the entire coding region of each gene was digested with BamHI/EcoRI and XhoI, and ligated into pMal-c2x vector (New England BioLab, Beverly, CA, USA) predigested with the same enzymes. The resulting plasmids were transformed into competent E. coli TB1 cells (Amersham Biosciences) and spread on Luria-Bertani (LB) agar plates containing 100 µg of ampicillin per ml. The expressions of recombinant proteins were induced by adding isopropyl-1-thio-β-D-galactopyranoside (IPTG) to a final concentration of 1 mM, and they were purified using an amylase affinity chromatography column (New England BioLab). The maltose-binding protein (MBP) was removed from the purified recombinant proteins using Factor Xa (New England BioLab) according to the manufacturer's instructions. The expressions and purities of the recombinant proteins were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE).

Oligonucleotide primers used for PCR amplifications of N. caninum antigen genes
Production of polyclonal antibodies
The polyclonal antisera directed against the whole lysates of N. caninum tachyzoites (NcWL) were produced by immunizing BALB/c mice with NcWL. Accordingly, mice were immunized intraperitoneally with NcWL (100 µg/mouse) emulsified with Freund's complete adjuvant. After primary immunization, animals were boosted 2 and 4 weeks later with the same antigen emulsified with Freund's incomplete adjuvant. Polyclonal antisera directed against the recombinant proteins, NcSAG1, NcSRS2, NcDG1, or NcDG2, were produced by immunizing 4 BALB/c mice with each purified protein. Mice were immunized intraperitoneally with purified protein (50 µg/mouse) emulsified with Freund's complete adjuvant, respectively. After primary immunization, animals were boosted 2 and 4 weeks later using the same protein emulsified with Freund's incomplete adjuvant. Sera were collected from immunized mice at 6 weeks after initial immunization, and immunoglobulin G was purified using a Protein G affinity column (Amersham Biosciences). Antibody titers were determined by enzyme-linked immunosorbent assay (ELISA) using relevant recombinant proteins as antigens.
Inhibitory effect of immunized sera on N. caninum invasion into host cells
To investigate the inhibitory effect of immunized sera on the invasion of N. caninum tachyzoites into host cells, in vitro inhibition assays were performed. Purified N. caninum tachyzoites (105) in PBS were mixed with two-fold stepwise dilutions (from 1:10 to 1:10,000) of each polyclonal antibody. Tachyzoites in PBS containing diluted normal mouse antibodies were used as negative controls. The mixtures were preincubated for 30 min at 37℃ and then added to Vero cells (2 × 105 cells) cultured in 8 well chamber slides. After 1 h incubation, the medium in each chamber was changed with fresh complete medium and incubated for 1 day at 37℃. Subsequently, infected parasites were differentiated by Giemsa staining and counted. Inhibition percent was expressed as [(% of infected cells in the negative control - % of infected cells after antibody treatment)/% of infected cells in the negative control] × 100. Neutralization titer was defined as the reciprocal of the antibody dilution that reduced parasite infection by 50% versus the negative control. All experiments were repeated at least three times.
Vaccination and challenge infection
Gerbils (Meriones unguiculatus, 5-6 weeks of age) were used for the challenge experiments. Uninfected control group animals (n = 23) were injected intraperitoneally with only PBS. The other experimental groups (n = 24 to 40/group) were injected intraperitoneally with NcWL or recombinant proteins. Animals in the first experimental group were injected with NcSAG1, NcSRS2, NcDG1, or NcDG2 at 100 µg/gerbil intraperitoneally, whereas animals in the second group were injected intraperitoneally the recombinant proteins in combinations. Gerbils were boosted with the same inoculums 2 weeks after the first injection. For parasite challenge, gerbils were inoculated intraperitoneally with live N. caninum tachyzoites (105) suspended in PBS, 1 week after boosting (3 weeks after the first injection). The protective efficacies of the vaccinations against infection were determined by comparing survival rates after challenge infection of the control group and each of the experimental groups using a Kruskal-Wallis non-parametric test and a distribution-free multiple comparisons method (Mutapi and Roddam, 2002). Significance was accepted at the p < 0.05 level.
RESULTS
Expression of N. caninum recombinant proteins and the production of polyclonal antibodies against the recombinant proteins
Four major antigens of N. caninum, NcSAG1, NcSRS2, NcDG1, or NcDG2, were expressed in E. coli and purified (Fig. 1A). Each purified recombinant antigen was immunized into mice to produce polyclonal antibodies against the proteins. Antibody reactivities were evaluated by Western blotting using each antibody and recombinant protein. Antibodies reacted specifically with their counterpart proteins and no cross reactivity was found (Fig. 1B). Antibody titers against the relevant antigens were < 1:10,000 by ELISA (data not shown).
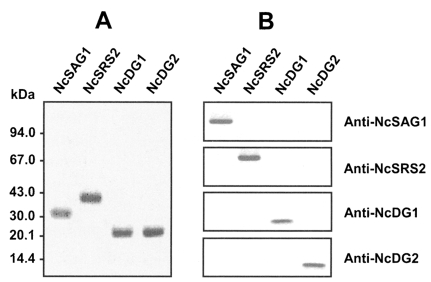
Expression, purification and Western blot analysis of N. caninum recombinant antigens. (A) Purification of N. caninum recombinant proteins. The expressions of recombinant proteins were induced by adding IPTG to a final concentration of 1 mM. Expressed proteins were purified by amylase affinity chromatography. The maltose-binding protein (MBP) was removed from the purified recombinant proteins by using Factor Xa and analyzed by 10% SDS-PAGE. The positions of molecular weight markers are indicated in kDa on the left side. (B) Western blot analysis. Purified proteins were separated by 12% SDS-PAGE, transferred to nitrocellulose membranes, and then probed with polyclonal antibody against each protein.
Inhibition of N. caninum invasion into host cells by treating with polyclonal antibodies
In vitro inhibition assays were performed to determine the inhibitory effect of each polyclonal antibody on the invasion of N. caninum tachyzoites into host cells (Fig. 2A). All antibodies had inhibitory activities ranging from 61.2 to 87.8%, as determined by parasite invasion into host cells at a 1:400 dilution. Neutralization titer, the reciprocal of antibody dilution giving 50% reduction of invasion of the parasite into host cells, of the antibodies were; 1:4,000, 1:900, 1:1,700, 1:2,500, and 1:600 for NcWL, NcSAG1, NcSRS2, NcDG1, and NcDG2, respectively (Fig. 2B). These neutralization titers indicate that NcSAG1, NcSRS2, NcDG1, or NcDG2 evoke antibody production that can effectively inhibit the invasion of N. caninum tachyzoites into host cells.
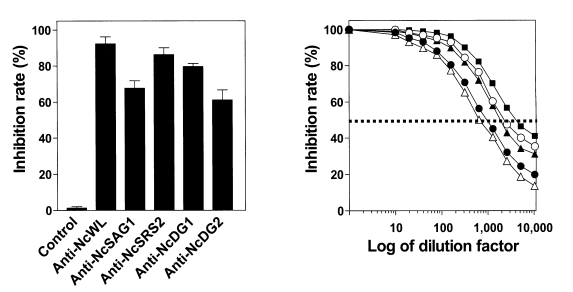
Inhibition of N. caninum tachyzoite invasion into host cells by polyclonal antibody treatment. (A) The inhibitory effect of polyclonal antibody treatment on N. caninum tachyzoite invasion into host cells. The inhibition rates shown represent means ± S.D. of triplicate experiments. (B) Determination of the neutralization titer of polyclonal antibody. Neutralization titer was defined as the reciprocal of antibody dilution that inhibited parasite invasion into host cells by 50%. Anti-NcWL (▪); anti-NcSAG1 (●); anti-NcSRS2 (▴); anti-NcDG1 (○); anti-NcDG2 (▵).
In vivo evaluation on the protective efficacies of vaccination of N. caninum recombinant antigens
It was difficult to evaluate the protective efficacies of recombinant antigens in mice, because most mice, e.g. BALB/c and C57BL/6, are resistant to N. caninum infection (Long et al., 1998). According to our preliminary study, gerbils are highly sensitive to N. caninum infection and all animals inoculated with 4 × 105 of N. caninum tachyzoites intraperitoneally died within 11 days with an LD50 value of 4 × 103 (data not shown). Therefore, in this study, we used gerbils as experimental animals to evaluate vaccination efficacy. Vaccination efficacy was evaluated by determining the survival durations and the survival rates of vaccinated gerbil groups after challenge infections. The protective efficacies of vaccinations of single- or multiple-recombinant antigens followed by challenge with N. caninum tachyzoites are summarized in Table 2. When negative controls were treated with N. caninum tachyzoites, 4 of the 23 gerbils (17.4%) survived. In animals vaccinated with NcWL, 17 of the 24 gerbils (70.8%) survived. All groups of gerbils vaccinated with single- or multiple-recombinant antigens also demonstrated meaningful levels of protection against the challenge infections. The survival rates of animals vaccinated with these recombinant antigens varied from 42.5 to 67.5%. The highest survival rate (67.5%) was obtained after vaccination with the recombinant antigens, NcSRS2 and NcDG1 in combination.
Survival rates of gerbils vaccinated with recombinant antigens and later challenged with N. caninum tachyzoites
DISCUSSION
Unlike the situation for other protozoan parasites, the development of an effective vaccine against neosporosis may be feasible because immunization with N. caninum tachyzoite lysates confers significant protection against infection in animal models (Liddell et al., 1999; Romero et al., 2004). Therefore, many studies have been undertaken to identify the antigens that elicit protective immunity against N. caninum infection in vitro and in vivo, in order to develop a reliable vaccine against neosporosis. In this study, we evaluated the protective efficacies of four N. caninum recombinant antigens against N. caninum infection in a gerbil model to identify optimal vaccine combination.
Two major surface proteins, NcSAG1 and NcSRS2, and two dense granule proteins, NcDG1 and NcDG2, of the parasite have been previously considered as effective vaccine candidate antigens that confer immunity against N. caninum infection (Innes et al., 2002). Specific antibodies against these proteins were found to inhibit the invasion of the parasite into host cells (Howe et al., 1998; Augustine et al., 1999; Nishikawa et al., 2001; Cannas et al., 2003). As was shown by these previous studies, all recombinant proteins tested in this study evoked antibody production that effectively inhibited parasite invasion into host cells. Inhibition rates ranged from 61.2 to 87.8% and anti-NcSRS2 and anti-NcDG1 showed much stronger inhibitory effects than anti-NcSAG1 and anti-NcDG2.
To evaluate the protective efficacy of recombinant antigen vaccination in vivo, we challenged gerbils vaccinated with recombinant antigens singly or in combination, and found that all vaccinated animals were significantly protected against the challenge infections, but that the NcSRS2 and NcDG1 combined vaccination had the greatest efficacy. This result was not in-line with our initial expectations, i.e., that a combination of all four antigens would probably be most efficacious. We do not know why some combination of all antigenic proteins did not have a maximal protective effect against experimental infection, though this may have been due to complexities associated with different immune reactions against each antigen in the host. Thus, further systematic immunological studies are required to determine why this combination of two antigens was found to have a stronger protective effect against experimental infection than some combination of all four antigens.
In conclusion, all the antigens tested in this study effectively induced antibody production that inhibited parasites infecting host cells in vitro. Moreover, the vaccination of these antigens singly or in combination into gerbils also revealed that they all meaningfully protected animals against experimental infection. The results of our in vitro and in vivo studies suggest that NcSRS2 and NcDG1 are the best combination for Neospora vaccine development.
Notes
This study was supported by a grant from the Agricultural R&D Promotion Center of the Republic of Korea (Project number 29909-3).