Enzymatic N-glycan analysis of 31 kDa molecule in plerocercoid of Spirometra mansoni (sparganum) and its antigenicity after chemical oxidation
Article information
Abstract
A highly specific antigenic protein of 31 kDa from plerocercoid of Spirometra mansoni (sparganum) was obtained by gelatin affinity and Mono Q anion-exchange column chromatography. The purified 31 kDa protein was subjected to N-glycan enzymatic digestion for structural analysis. The relative electrophoretic mobility was analyzed by SDS-PAGE, before and after digestion. On SDS-PAGE after enzymatic digestion, the 31 kDa protein showed a molecular shift of approximately 2 kDa, which indicated the possession of complex N-linked oligosaccharides (N-glycosidase F sensitive) but not of high-mannose oligosaccharides (endo-beta-N-acetylglucosaminidase H, non-sensitive). Chemically periodated 31 kDa protein showed statistically non-significant changes with human sparganosis sera by enzyme linked immunosorbent assay (ELISA). Therefore, the dominant epitopes of the 31 kDa molecule in human sparganosis were found to be mainly polypeptide, while N-glycans of the antigenic molecule in sparganum was minimal in anti-carbohydrate antibody production.
INTRODUCTION
Human sparganosis is a disease caused by the ingestion of plerocercoid present in snakes or procercoid present in drinking spring water in Korea (Cho et al., 1975). Serologic diagnosis of human sparganosis is available using enzyme-linked immunosorbent assay (ELISA) (Kim et al. 1984) and immunoblotting (Choi et al., 1988).
Parasite infection provokes an immune responses to glycoproteins, mainly directed against glycoconjugates. In schistosomiasis, humoral immune response against several schistosomal glycoconjugates in infected animals has been found (Nyame et al., 1999; Nyame et al., 2000; Van Die et al., 1999). The oligosaccharide structures of pathogens are important in the first contact with the host innate immune system. In Leishmania sp., cell surface oligosaccharides in promastigotes play a role in the macrophage infection in mouse leishmaniasis (Channon et al., 1984; Blackwell et al., 1985). Moreover, oligosaccharides may also provoke an antibody response and serve as a target for specific antibodies. Protective monoclonal antibodies were developed against the excretory-secretory antigens from Trichinella spiralis muscle larvae - TSL1 (Ellis et al., 1994).
The 31 kDa molecule in the excretory-secretory product (Cho et al., 1992) known as a protease in crude extracts showed a highly specific and sensitive reaction in human sparganosis (Choi et al., 1988). Antigenic molecules in parasitic infection appeared to be distributed mainly intracellular or on the surfaces as a glycoprotein. The 31 kDa antigenic molecule in sparganum is localized in the tegument, especially in the syncytial tegument and tegumental cells (Kim et al., 1992). This study examined the carbohydrate moieties of purified 31 kDa molecule from sparganum by enzymatic deglycosylation and the effects of carbohydrate epitopes in its antigenicity in human sparganosis.
MATERIALS AND METHODS
Worms and crude extracts
Sparganum was collected from the naturally infected terrestrial snake, Elaphe rufodorsata. Collected worms were washed in physiological saline 3 times, and homogenized by a Teflon-pestle homogenizer with five times volume of 20mM Tris-HCl buffer (pH 7.4). The product was centrifuged at 15,000 rpm for 1 hr, and the resulting supernatant was used as a crude extract, and stored at -70℃ until experimental use after the protein concentration was measured by the Lowry method.
Purification of 31 kDa molecule
The 31 kDa protein was purified by gelatin affinity chromatography (Kong et al., 1991) and Mono Q HR 5/5 column using the AKTA FPLC system (Amersham Pharmacia Biotech, Piscataway, NJ, USA). Crude extracts equilibrated with 20 mM Tris-HCl (pH 7.4) were loaded on a Mono Q column. The column was washed with the buffer and non-specific binding proteins were eluted at a flow rate of 0.5 ml/min. The adsorbed proteins were eluted with 1 M NaCl in the same buffer by stepwise gradient fashion. Purified protein was monitored by 7.5-15% gradient SDSPAGE. Purified fraction was dialyzed with 20 mM Tris-HCl (pH 7.4) and stored at -70℃.
Enzymatic digestion
N-glycosidase F (PNGase F) from Flavobacterium meningosepticum (Roche, Mannheim, Germany), and endo-beta-N-acetylglucosaminidase H (Endo H) from Streptomysecs plicatus (Roche, Mannheim, Germany) were used in enzymatic deglycosylation experiments. The deglycosylation conditions for enzymes on purified 31 kDa antigenic molecule by those enzymes were adopted from manufacturer's instruction. Fetuin (Sigma, St. Louis, MO, USA) was used as a positive control in PNGase F digestion. After enzymatic treatment, all antigenic protein was separated by 7.5~15% gradient SDS-PAGE and stained with Coomassie brilliant R-250 solution to determine any shift in the 31 kDa molecule.
Chemical oxidation with sodium meta-periodate and ELISA
Optimal chemical oxidation of glycoprotein with sodium meta-periodate has been known that the hexose ring can be linearized and destroy carbohydrate related immunological reactivities. However, it does not alter the structure of the polypeptide chains. (Eylar and Jaenloz, 1962; Miller-Podraza and Fishman, 1982). To determine the optimal concentration (50 mM) of sodium meta-periodate (Sigma St. Louis, MO, USA), chemically treated 20 µg crude extracts (final 5~200 mM) were separated on 7.5~15% SDS-PAGE. ELISA method was performed as described previously (Schallig and van Leeuwen, 1996; Woodward et al., 1985) with minor modification. In ELISA, each native crude extract (5 µg/ml) and the purified 31 kDa molecule (1 mg/m) were coated on 96-well plates as an antigen. Surgically confirmed human sparganosis sera were used in these experiments (Kong et al., 1994). Absorbance was measured in triplicate at 490 nm and expressed as the mean ± SD.
RESULTS
Purified 31 kDa antigenic protein was demonstrated on 7.5-15% SDS-PAGE (Fig. 1A) and its broad band means that it might contain the carbohydrate modification. The 31 kDa molecule with PNGase F treatment showed a reduction of about 2 kDa, leaving a 29 kDa molecule which was sensitive to PNGase F. As shown in Fig. 1B, there was no change in molecular weight following digestion with Endo H. Based on these findings, it was concluded that the observed decrease in the molecular weight of the 31 kDa molecule was likely a result of the specific removal of complex N-linked oligosaccharide structure from N-glycans of the 31 kDa antigenic protein.
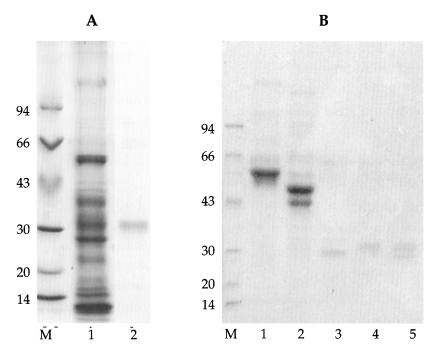
A. Purified 31 kDa antigenic molecule of sparganum on 7.5-15% gradient SDS-PAGE. 1 crude extracts, 2 purified 31 kDa protein. B. lane 1, fetuin; lane 2, fetuin treated with PNGase F; lane 3, 31 kDa protein with PNGase F; lane 4, 31 kDa protein; lane 5, 31 kDa protein treated with Endo H. Note the molecular shift on PNGase F treatment and not on Endo H digestion (lane 5). (M; molecular marker, kDa)
To investigate the antigenic role of carbohydrate moieties of the purified 31 kDa molecule, native and chemically meta-periodated proteins were evaluated using sparganosis sera by ELISA. Although statistically non-significant in comparison with the chemical direct effects on polypeptides of the 31 kDa protein, minimal reductions of absorbance in both sodium meta-periodate treated crude and purified 31 kDa molecule are observed in Table 1. These results suggest that the 31 kDa molecule has dominant epitopes of polypeptides in human sparganosis. In addition, in Western blot, we could not discriminate the effects of carbohydrates on anti-31 kDa antibody production (data not shown).

Absorbance changes after sodium meta-periodate treatment on crude extracts and purified 31 kDa protein measured by ELISA
DISCUSSION
The findings of the present study are similar with those of March et al. (1991) who showed no significant differences in antigenicity of PNGase F and Endo H treated Antigen 5 of Echinococcus granulosus, which is the dominant polypeptide epitopes in Western blot. The immunogenicity of some parasite glycans may be due to a biochemical structure which differs from that of the mammalian host glycans. This suggests that the glycosylation pathways between host and parasite may be different due to their evolutionary distances (Ferguson and Homans, 1988).
Antibody responses against sparganum infection may be strong due to its lifelong larval stage in the human. Therefore, some modifications may provoke carbohydrate epitopes of antigenic molecules, especially in surface or secretory proteins of sparganum. Even though the secreted 31 kDa protein has long carbohydrate chains, the antigenicity of the meta-periodated 31 kDa protein showed a non-significant effect on anti-carbohydrate antibody production. Although the biological significance of the N-glycans structure (complex N-linked oligosaccharide) in the 31 kDa antigenic protein is unknown, it the precise role of carbohydrate epitopes in the immunogenicity of the glycoproteins remains to be elucidated. Therefore, further studies are required to rule out the presence of 'hidden' carbohydrates in sparganum from the host immune system or O-linked oligosaccharides in sparganum antigens.
Notes
This work was supported by a Korea Research Foundation grant (KRF-2001-041-F00077).