Seo, Araujo, Reinhard, Chai, and Shin: Paleoparasitological Studies on Mummies of the Joseon Dynasty, Korea
Abstract
Paleoparasitology is the application of conventional or molecular investigative techniques to archeological samples in order to reveal parasitic infection patterns among past populations. Although pioneering studies already have reported key paleoparasitological findings around the world, the same sorts of studies had not, until very recently, been conducted in sufficient numbers in Korea. Mummified remains of individuals dating to the Korean Joseon Dynasty actually have proved very meaningful to concerned researchers, owing particularly to their superb preservation status, which makes them ideal subjects for paleoparasitological studies. Over the past several years, our study series on Korean mummies has yielded very pertinent data on parasitic infection patterns prevailing among certain Joseon Dynasty populations. In this short review, we summarized the findings and achievements of our recent paleoparasitological examinations of Joseon mummies and discussed about the prospects for future research in this vein.
Key words: paleoparasitology, parasite, mummy, Joseon Dynasty, Korea
INTRODUCTION
Paleoparasitology is the study of parasites found in ancient remains and the tracing, on that basis, of the history of host-parasite relationships [ 1, 2]. It began in the early 20th century in Egypt [ 3], where Sir Marc Armand Ruffer developed a special rehydration technique applicable to mummified tissues and prepared histological slides suitable for their examination [ 4]. Over the succeeding several decades, paleoparasitology continued to develop all over the world, as experts examined sediments or coprolites obtained from archaeological sites, discovering therein ancient parasite eggs by conventional or molecular techniques. In Korea, such samples are now the focus of interdisciplinary collaboration, most notably between archaeologists and parasitologists seeking to trace and delineate past parasitic infection patterns. Some of the fruits of this labor will be reviewed briefly here.
BRIEF HISTORY OF PALEOPARASITOLOGY IN KOREA
The first paleoparasitological investigation in Korea was carried out on Ascaris and Trichuris eggs discovered in wetland soil samples obtained in Shinchang-Dong [ 5]. Although this was not a full-scale survey, it was nonetheless meaningful for being the first study of its kind in Korea. Several years later, another Korean paleoparasitological investigation was conducted, this one on archaeological soil samples dating to the Bronze Age (2000-1000 BC), the Three-Kingdoms Period (100 BC-650 AD), the Unified Silla Dynasty (668-935 AD) and the Joseon Dynasty (1392-1910) [ 6]. As in the Shinchang-Dong case, nematode (i.e. Ascaris and Trichuris) and trematode (i.e. Clonorchis) eggs were discovered. This study was remarkable, though, for representing the first actual Korean archaeological-parasitological collaboration by which parasitic infection patterns for a different historical era could be revealed [ 6]. Over time, mummies discovered in tombs dating to the Joseon Dynasty stood out as especially relevant and valuable to paleoparasitology. These tombs in fact had been constructed in significant numbers throughout most of the Korean peninsula, mainly during the latter half of the period (16th-18th century). According to historical texts, they were very strongly built tombs. In fact,
...since lime, sand, and red clay (filled around the coffin) became hardened like a stone after a long while, the coffin could be protected from the possible intrusions of insects or even grave robbers.... [ 7]
Fortunately enough for archaeologists, these Joseon tombs allowed for superb preservation of the cultural artifacts and human remains they contained, though the exact mechanism of preservation was not fully, and remains imperfectly, understood. The former has proved instrumental in revealing detailed aspects of Joseon society [ 8]. As for the latter, there have been some exceptionally well preserved Joseon mummies that have yielded particularly valuable evidence to investigators ( Fig. 1A). Coprolites for example, generally shown on computed tomography (CT) images as well-demarcated masses in the intestinal cavity [ 8], have been collected from Joseon mummies in the course of endoscopic examinations or by direct dissection, and have proved ideal for the broad purposes of paleoparasitological studies in Korea ( Fig. 1A, B). In these samples, various ancient parasite species have been found, including Trichuris trichiura and Ascaris lumbricoides [ 8, 9]. These findings, indeed, have begun to paint pictures of parasitic infection patterns prevailing during the Joseon Dynasty. In the course of sample collection, every effort has been made to minimize false-positivity due to contamination: investigators wore protective gloves, head cap, gown, and mask ( Fig. 2A-C), and surface soils from the given archaeological site were always applied as negative controls. As for findings, our own data accumulated over the past several years, and especially our accurate predictions of the presence of parasite eggs in a number of sample sets, indicates a close relationship between the preservation status of certain types of artifacts or remains (e.g., clothes, hair, or brain tissue) and that of ancient parasite eggs discovered in the same tombs [ 10]. Microscopic examinations of samples (typically rehydrated in 0.5% trisodium phosphate solution) have been the standard investigative modality [ 11, 12]. What has really captured the imagination of scientists, even of archaeologists, over the past several decades, however, is DNA analysis [ 13, 14, 15]. DNA studies of ancient parasites including Trypanosoma cruzi, Plasmodium sp., Ascaris sp., T. trichiura, Enterobius vermicularis, and Clonorchis sinensis have been numerously and widely reported [ 13, 16, 17, 18, 19, 20, 21, 22]. Inspired by these successes, our research team has performed ancient DNA (aDNA) analyses on many of the parasite egg specimens we have obtained from Joseon mummies [ 23, 24].
PALEOPARASITOLOGICAL STUDIES ON JOSEON MUMMIES
The availability of coprolite samples from Joseon Dynasty mummies has proved very fortunate indeed. Due to their great preservation quality, we were able to collect invaluable data on the patterns of parasitic infections affecting certain Joseon societies ( Fig. 6A). The first-ever paleoparasitological study on a Joseon population was based on a child mummy discovered in 2001 ( Fig. 2D) in a 16th to 17th century Joseon tomb unearthed in Yangju, Gyonggi-do ( Fig. 6A, No. 13). Endoscopy revealed the presence of coprolites, samples of which showed, under scanning electron microscopy (SEM), T. trichiura, A. lumbricoides and C. sinensis eggs still perfectly intact even after several hundreds of years [ 9, 25] ( Fig. 3E, F). In another investigation, this one involving a 17th century Joseon tomb unearthed in Gongju ( Fig. 6A, No. 15), the sample's preservation was so perfect that even larvae of various species, in addition to eggs, were detected [ 26]. In the course of our paleoparasitological study series, we have found ancient parasite eggs and larvae representing A. lumbricoides, T. trichiura, C. sinensis, Paragonimus westermani, Metagonimus yokogawai, Gymnophalloides seoi, Strongyloides stercoralis, and Trichostrongylus spp. ( Figs. 3, 4, 5) [ 8, 9, 10, 26, 27, 28, 29]. These results were summarized in Table 1. These and other, related data have answered many questions concerning historical parasitic infection patterns in Korea. Among our most noteworthy findings was that the infection prevalence of nematode parasites in Joseon samples does not differ so much from those of mid-20th century Korea. In fact, the overall positive rates for A. lumbricoides and T. trichiura in our Joseon data were 50.0% (9/18) and 77.8% (14/18), respectively, compared with 1969 national survey results of 58.2% ( A. lumbricoides) and 74.5% ( T. trichiura) [ 30]. Notwithstanding this general consistency with the national survey data of 1969, there were some unique findings represented in our data. The Joseon samples contained some parasite species not readily identifiable, such as E. vermicularis, hookworms, and Taenia spp. To date, there have been only 2 reports of E. vermicularis eggs in archaeological samples: one on Roman latrines, and the other on an Egyptian mummy [ 31, 32]. Ours, then, is the first such report for any East Asian country [ 33]. In feces remnant in the colon of a female Joseon mummy from Dangjin-gun ( Fig. 6A, No. 8), E. vermicularis along with P. westermani and A. lumbricoides eggs were identified under microscopy. Another rare finding from our Joseon samples was human taeniasis. Its infection prevalence among Koreans reportedly was only 1.9% in 1971 [ 34], and it had not been found in any archaeologically obtained Korean samples other than our 17th century Joseon case [ 35]. We will need to examine additional samples before we are able to confirm this rarity or, if confirmed, to determine the corresponding infection route it is relatable to the dietary habits of the Joseon people. In Joseon mummy coprolites we have observed many trematode eggs ( Table 1), among which those of C. sinensis eggs have been very common [ 9, 20, 27, 36]. Considering that C. sinensis infection can be transmitted via ingestion of raw or undercooked freshwater fish harboring the infectious metacercariae [ 37, 38], we speculated that this Korean culinary tradition might in fact have been practiced by pre-20th century Koreans more than that had previously been supposed. Another trematode parasite frequently found in Joseon mummy coprolite samples has been P. westermani [ 10, 26, 29, 33, 39]. These many cases of P. westermani eggs show that this type of infection might not have been rare in Joseon populations. Infected individuals might have ingested crabs or crayfish in a raw or undercooked state, especially considering the fact that freshwater crustaceans are the second intermediate hosts of Paragonimus spp. Among our cases of paragonimiasis, the most impressive and, incidentally, the first-ever archaeological-sample-based case of ectopic paragonimiasis reported anywhere in the world, is a ca 17th century female mummy discovered in 2009 ( Fig. 6A, No. 4) [ 29]. According to the extant historical texts, she was likely to have lived in Hadong sometime between the late 1500s and early 1600s. On microscopic examinations, many Paragonimus eggs were found in the liver, lung, intestinal tissues, and feces ( Fig. 5). Other very interesting findings were G. seoi eggs in the coprolites of another female mummy discovered in Hadong ( Fig. 6A, No. 5) [ 27]. This was a very unexpected and illuminating discovery, as there had been no previous reports of G. seoi eggs anywhere in Hadong-gun (county), the area in which the mummified individual had lived. The first report of G. seoi infection was made in 1993, concerning a Korean woman residing on an island off the southwest coast of Korea; and in fact, a 2001 nationwide survey established that human G. seoi infection was confined to that and other small islands or counties in the southwestern corner of the country [ 40, 41]. In this light, we suspected that G. seoi infection might have contracted from what had been a wider coastal distribution to a much more restricted region of the Korean peninsula [ 27] ( Fig. 6B). This, our contraction theory of G. seoi infection, though convincing to us, required more than just a single mummy case from a non-endemic area to be considered dispositive. Fortunately enough, we did discover, in Sapgyo, a 16th century Joseon mummy ( Fig. 6A, No. 3) in which, along with C. sinensis, M. yokogawai, and T. trichiura, G. seoi eggs were evident [ 39]. In one of our other analyses, this one of ancient Ascaris eggs from another Joseon specimen, we were successful in extracting and sequencing Ascaris aDNA [ 23]. However, the Ascaris 18S rRNA gene sequence in our study was more similar to A. suum than to A. lumbricoides, though the samples were collected from Joseon human individual remains during anthropological investigations. As in the previous relevant studies, we speculated, therefore, that aDNA analysis of Ascaris is inadequate for confirmation of A. lumbricoides [ 23, 42, 43, 44]. In our PCR-based aDNA analysis of the T. trichiura SSU rRNA gene, we could obtain a sequence 100% homologous to that of T. trichiura, which is also distinct from that of T. suis (97%) and T. muris (91%) ( Table 2) [ 24]. We have applied aDNA analysis also to the study of trematode eggs, specifically Paragonimus eggs collected from a 17th century Korean female mummy ( Fig. 6A, No. 4) [ 29]. The extracted ITS 2 gene sequence showed 100% homology to those of modern P. westermani DNA reported from Korea and Japan, which collective results constituted a genetic cluster distinct from other South Asian P. westermani. Subsequently, aDNA sequences of C. sinensis were analyzed. Although ancient C. sinensis eggs have frequently been observed in Korean archaeological samples [ 9, 20, 27, 36], reported analyses of C. sinensis aDNA have been very scarce. In one of our studies on a Joseon mummy, the presence of C. sinensis eggs was confirmed by microscopic observations, and C. sinensis aDNA was successfully extracted. A PCR-based aDNA analysis was then performed using primer sets for nuclear ribosomal internal transcribed spacers 1 (ITS1) and 2 (ITS2), and mitochondrial cytochrome c oxidase 1 (CO1) genes. The obtained sequences, significantly, were 100% homologous to the contemporary C. sinensis gene sequences reported from Korea and other East Asian countries [ 36] ( Table 2).
PROSPECTS OF PALEOPARASITOLOGY IN KOREA
For the past several years, we have studied Korean mummies and their paleoparasitological samples, which subsequently have been maintained in the Joseon Dynasty Human Remains Collection (JDHRC) of Seoul National University as the fruit of our collaboration with archaeologists. Although our paleoparasitological studies in Korea have provided important information on parasitic infection patterns, we must admit that most of the credit is owed to the discovery of well-preserved Joseon mummies. Our estimations of parasite prevalence, moreover, are based on samples representing relatively limited time, regional, and socio-economic spans. For instance, we should acknowledge that most of the Joseon mummies we have examined were individuals who belonged to the ruling classes of society. According to historians, the unique tombs ( Hoegwakmyo tombs) in which Korean mummies have been discovered were designed and constructed exclusively for such people. This means that the mummies are likely to have been landowners, scholars, or courtiers [ 45]. In a sense then, the coprolites of Korean mummies have helped to reveal the parasitic infection status not so much of a certain Joseon society in general but rather of the highest social strata of that society. In future studies, therefore, our scope will be extended to types of paleoparasitological samples other than those from Korean mummies. A great deal of data from various origins will likely prove fertile soil for the development of comprehensive theories on past parasitic infection patterns. Many questions and issues remain, the answering and resolving of which will occupy current work and guide subsequent future research. In our upcoming studies, we must try to find some types of helminth eggs that heretofore have not been considered. As already explained, some parasite species, with very few exceptions, have not been found in coprolite samples from Joseon mummies, despite their high reported prevalence in recent national survey reports. Examples of these are E. vermicularis, hookworms, and Taenia. As for protozoan cysts, those of Entamoeba histolytica or Giardia lamblia have gone undiscovered as well. Many intensive and detailed studies are still needed in order to identify certain parasite eggs, larvae, cysts, and oocysts rarely discovered in previous studies.
Also, whereas our previous aDNA studies served to confirm the academic significance of combining PCR-based molecular data with microscopic findings on ancient parasite eggs, such analysis will have to be extended to much deeper levels of inquiry and detail. We admit, for example, that aDNA study focusing more on phylogenetic analysis of ancient parasites over wider geographical and temporal ranges is required in order to broaden our understanding of the genetic differences among specific parasite species [ 46]. In conclusion, paleoparasitology is now on the advance, and growing as a field worldwide. Expanding cooperation among parasitologists, archaeologists and, more recently, palaeontologists, provides the comprehensive context in which the parasitic infection patterns of the past and, thereby, the origins and evolution of infectious diseases worldwide, can be fully understood. In Korea, paleoparasitological approaches to the study of archaeological samples are really just beginning. Even so, we expect that our forthcoming analyses finally will move even closer to the goal of revealing the parasitological infection patterns of Joseon society. Indeed, interpreting our data outcomes from the global parasitological perspectives will help to reconstruct a history of pre-modern parasitic infections for the entire Asian continent.
National Research Foundation of Korea2013R1A1A2009688
Brazilian agencies CAPES/Ciência sem Fronteiras
ACKNOWLEDGMENTS
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2013R1A1A2009688). Adauto Araújo's and Karl Reinhard's research is also funded by the Brazilian agencies CAPES/Ciência sem Fronteiras, CNPq, and FAPERJ.
Conflict of interest
We declare that we do not have any conflict of interest related to this study.
REFERENCES
1. Ferreira LF, Araújo A, Confalonieri U. Subsidios para a Paleoparasitologia do Brasil I - Parasitos encontrados em coprólitos no município de Unai, Minas Gerais. Congresso Brasileiro De Parasitologia. 2. Resumos. Campinas, SP. 1979, p 56.
2. Ferreira LF, Reinhard K, Araújo A. Fundamentos da Paleoparasitologia. Rio de Janeiro, Brazil. Editora Fiocruz. 2011, p 484.
3. Ruffer MA. Note on the presence of Bilharzia haematobia in Egyptian mummies of the twentieth dynasty (1250-1000). Br Med J 1910;1:16. PMID: 20764829.  4. Dittmar K, Araújo A, Reinhard KJ. The study of parasites through time: archaeoparasitology and paleoparasitology. In Grauer AL ed, A companion to paleopathology. Oxford, UK. Blackwell Publishing Co. 2012, pp 170-190.
5. Kanehara M, Kanehara M. Analysis of Shinchangdong Jeosupji Hwabun and parasite eggs. Gwangju Shinchangdong Jeosupji Yujeok. Gwangju, Korea. National Museum of Gwangju. 1997, Vol. I.
6. Han ET, Guk SM, Kim JL, Jeong HJ, Kim SN, Chai JY. Detection of parasite eggs from archaeological excavations in the Republic of Korea. Mem Inst Oswaldo Cruz 2003;98(Suppl 1):123-126. PMID: 12687771. 7. Chu H. Im MH, translator. Jujagarye. Seoul, Korea. Yemunseowon. 1999.
8. Lee IS, Lee EJ, Park JB, Baek SH, Oh CS, Lee SD, Kim YS, Bok GD, Hong JW, Lim DS, Shin MH, Seo M, Shin DH. Acute traumatic death of a 17th century general based on examination of mummified remains found in Korea. Ann Anat 2009;191:309-320. PMID: 19345566.  9. Seo M, Guk SM, Kim J, Chai JY, Bok GD, Park SS, Oh CS, Kim MJ, Yil YS, Shin MH, Kang IU, Shin DH. Paleoparasitological report on the stool from a medieval child mummy in Yangju, Korea. J Parasitol 2007;93:589-592. PMID: 17626351. 10. Seo M, Oh CS, Chai JY, Lee SJ, Park JB, Lee BH, Park JH, Cho GH, Hong DW, Park HU, Shin DH. The influence of differential burial preservation on the recovery of parasite eggs in soil samples from Korean medieval tombs. J Parasitol 2010;96:366-370. PMID: 19954260. 11. Callen EO, Cameron TWM. A pre-historic diet revealed in coprolites. New Scientist 1960;8:35-40.
12. Van Cleave HJ, Ross JA. A method for reclaiming dried zoological specimens. Science 1947;105:318. PMID: 17740567. 13. Iñiguez AM, Reinhard K, Carvalho Gonçalves ML, Ferreira LF, Araújo A, Paulo Vicente AC. SL1 RNA gene recovery from Enterobius vermicularis ancient DNA in pre-Columbian human coprolites. Int J Parasitol 2006;36:1419-1425. PMID: 16950265. 14. Martinez EM, Correia JA, Villela EV, Duarte AN, Ferreira LF, Bello AR. Random amplified polymorphic DNA analysis of DNA extracted from Trichuris trichiura (Linnaeus, 1771) eggs and its prospective application to paleoparasitological studies. Mem Inst Oswaldo Cruz 2003;98(Suppl 1):59-62. PMID: 12687764. 15. Marota I, Rollo F. Molecular paleontology. Cell Mol Life Sci 2002;59:97-111. PMID: 11846037. 16. Aufderheide AC, Salo W, Madden M, Streitz J, Buikstra J, Guhl F, Arriaza B, Renier C, Wittmers LE Jr, Fornaciari G, Allison M. A 9,000-year record of Chagas' disease. Proc Natl Acad Sci 2004;101:2034-2039. PMID: 14766963.  17. Liu WQ, Liu J, Zhang JH, Long XC, Lei JH, Li YL. Comparison of ancient and modern Clonorchis sinensis based on ITS1 and ITS2 sequences. Acta Trop 2007;101:91-94. PMID: 17276379. 18. Iñiguez AM, Reinhard KJ, Araújo A, Ferreira LF, Vicente ACP. Enterobius vermicularis: ancient DNA from North and South American human coprolites. Mem Inst Oswaldo Cruz 2003;98(Suppl 1):67-69. PMID: 12687766. 19. Guhl F, Jaramillo C, Vallejo GA, Yockteng R, Cárdenas-Arroyo F, Fornaciari G, Arriaza B, Aufderheide AC. Isolation of Trypanosoma cruzi DNA in 4,000-year-old mummified human tissue from northern Chile. Am J Phys Anthropol 1999;108:401-407. PMID: 10229385. 20. Sallares R, Gomzi S. Biomolecular archaeology of malaria. Ancient Biomol 2001;3:195-213.
21. Loreille O, Roumat E, Verneau O, Bouchet F, Hänni C. Ancient DNA from Ascaris: extraction amplification and sequences from eggs collected in coprolites. Int J Parasitol 2001;31:1101-1106. PMID: 11429174. 22. Leles D, Araújo A, Ferreira LF, Vicente ACP, Iñiguez AM. Molecular paleoparasitological diagnosis of Ascaris sp. from coprolites: new scenery of ascariasis in pre-Columbian South America times. Mem Inst Oswaldo Cruz 2008;103:106-108. PMID: 18327505. 23. Oh CS, Seo M, Lim NJ, Lee SJ, Lee EJ, Lee SD, Shin DH. Paleoparasitological report on Ascaris aDNA from an ancient East Asian sample. Mem Inst Oswaldo Cruz 2010;105:225-228. PMID: 20428686. 24. Oh CS, Seo M, Chai JY, Lee SJ, Kim MJ, Park JB, Shin DH. Amplification and sequencing of Trichuris trichiura ancient DNA extracted from archaeological sediments. J Archaeol Sci 2010;37:1269-1273. 25. Shin DH, Lim DS, Choi KJ, Oh CS, Kim MJ, Lee IS, Kim SB, Shin JE, Bok GD, Chai JY, Seo M. Scanning electron microscope study of ancient parasite eggs recovered from Korean mummies of the Joseon Dynasty. J Parasitol 2009;95:137-145. PMID: 18601576. 26. Shin DH, Chai JY, Park EA, Lee W, Lee H, Lee JS, Choi YM, Koh BJ, Park JB, Oh CS, Bok GD, Kim WL, Lee E, Lee EJ, Seo M. Finding ancient parasite larvae in a sample from a male living in late 17th century Korea. J Parasitol 2009;95:768-771. PMID: 19071966. 27. Seo M, Shin DH, Guk SM, Oh CS, Lee EJ, Shin MH, Kim MJ, Lee SD, Kim YS, Yi YS, Spigelman M, Chai JY. Gymnophalloides seoi eggs from the stool of a 17th century female mummy found in Hadong, Republic of Korea. J Parasitol 2008;94:467-472. PMID: 18564747. 28. Seo M, Oh CS, Shin DH. Parasite Eggs found in Korean Mummies. Yearbook of Mummy Studies. Seoul, Korea. 2011, Vol. 1:pp 75-81.
29. Shin DH, Oh CS, Lee SJ, Lee EJ, Yim SG, Kim MJ, Kim YS, Lee SD, Lee YS, Seo M. Ectopic paragonimiasis from 400 year old female mummy of Korea. J Archaeol Sci 2012;39:1103-1110. 30. Seo BS, Rim HJ, Loh IK, Lee SH, Cho SY, Park SC, Bae JW, Kim JH, Lee JS, Koo BY, Kim KS. Study on the status of helminthic infections in Koreans. Korean J Parasitol 1969;7:53-70. 31. Herrmann B. Parasitologisch-epidemiologische auswertungen mittelalterlicher kloaken. Z Archaol Mittelalters 1985;13:131-161.
32. Horne PDJ. First evidence of enterobiasis in ancient Egypt. J Parasitol 2002;88:1019-1021. PMID: 12435149. 33. Shin DH, Oh CS, Chai JY, Lee HJ, Seo M. Enterobius vermicularis eggs discovered in coprolites from a medieval Korean mummy. Korean J Parasitol 2011;49:323-326. PMID: 22072838. 34. Eom KS, Rim HJ. Epidemiological understanding of Taenia tapeworm infections with special reference to Taenia asiatica in Korea. Korean J Parasitol 2001;39:267-283. PMID: 11775327. 35. Lee HJ, Shin DH, Seo M. Discovery of taeniid eggs from a 17th century tomb in Korea. Korean J Parasitol 2011;49:327-329. PMID: 22072839. 36. Shin DH, Oh CS, Lee HJ, Chai JY, Lee SJ, Hong DW, Lee SD, Seo M. Ancient DNA analysis on Clonorchis sinensis eggs remained in samples from medieval Korean mummy. J Archaeol Sci 2013;40:211-216. 37. Kang SY, Ahn IY, Park CY, Chung YB, Hong ST, Kong Y, Cho SY, Hong SJ. Clonorchis sinensis: molecular cloning and characterization of 28-kDa glutathione S-transferase. Exp Parasitol 2001;97:186-195. PMID: 11384162. 38. Yu SH, Masanori K, Li XM, Xu LQ, Lan CG, Lin R. Epidemiological investigation on Clonorchis sinensis in human population in an area of south China. Jpn J Infect Dis 2003;56:168-171. PMID: 14583642. 39. Shin DH, Oh CS, Chai JY, Ji MJ, Lee HJ, Seo M. Sixteenth century Gymnophalloides seoi infection on the coast of the Korean Peninsula. J Parasitol 2012;98:1283-1286. PMID: 22524319. 40. Lee SH, Chai JY, Hong ST. Gymnophalloides seoi n. sp. (Digenea: Gymnophallidae), the first report of human infection by a gymnophallid. J Parasitol 1993;79:677-680. PMID: 8410538. 41. Chai JY, Park JH, Han ET, Shin EH, Kim JL, Hong KS, Rim HJ, Lee SH. A nationwide survey of the prevalence of human Gymnophalloides seoi infection on western and southern coastal islands in the Republic of Korea. Korean J Parasitol 2001;39:23-30. PMID: 11301587. 42. Leles D, Araújo A, Vicente AC, Iñiguez AM. ITS1 intra-individual variability of Ascaris isolates from Brazil. Parasitol Int 2010;59:93-96. PMID: 19819347. 43. Anderson TJC. The dangers of using single locus markers in parasite epidemiology: Ascaris as a case study. Trends Parasitol 2001;17:183-188. PMID: 11282508. 44. Peng W, Yuan K, Hu M, Zhou X, Gasser RB. Mutation scanning-coupled analysis of haplotypic variability in mitochondrial DNA regions reveals low gene flow between human and porcine Ascaris in endemic regions of China. Electrophoresis 2005;26:4317-4326. PMID: 16287175. 45. Shin MH, Yi YS, Bok GD, Lee EJ, Spigelman M, Park JB, Min SR, Shin DH. How did mummification occur in bodies buried in tombs with a lime soil mixture barrier during the Joseon Dynasty in Korea. In Pena PA, Martin RM, Rodriguez AR eds, Mummies and Science. Santa Cruz de Tenerife, Spain. World Mummies Research. 2008, pp 105-113.
46. Martinsen ES, Perkins SL, Schall JJ. A three-genome phylogeny of malaria parasites ( Plasmodium and closely related genera): evolution of life-history traits and host switches. Mol Phylogenet Evol 2008;47:261-273. PMID: 18248741.
Fig. 1
Parasitological sampling from Korean mummy. (A) Making a hole for endoscopic exploration for Korean mummy. Coprolites could be collected during endoscopic examination. (B) Very well preserved feces could be obtained during the dissection of Gangneeung mummy. 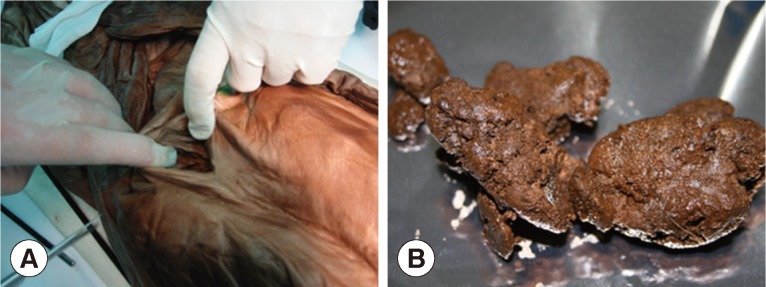
Fig. 2
(A) False-positivity due to the contamination of sample could be ruled out by examination of the archaeological site's surface soils. The negative control sample could be obtained from the surface soil (black dot) adjacent to the tomb. (B) When the parasitological samples were collected from geological strata, negative control could be obtained from the nearby surface soil (black dot). (C) Joseon mummies discovered tightly wrapped in layers of clothing and other textiles. Researchers wearing sterilized gowns, masks, head caps, and gloves tried to remove the textile before paleoparasitological sampling. (D) The paleoparasitological study on a child mummy of Joseon period, found in Yangju-gun. 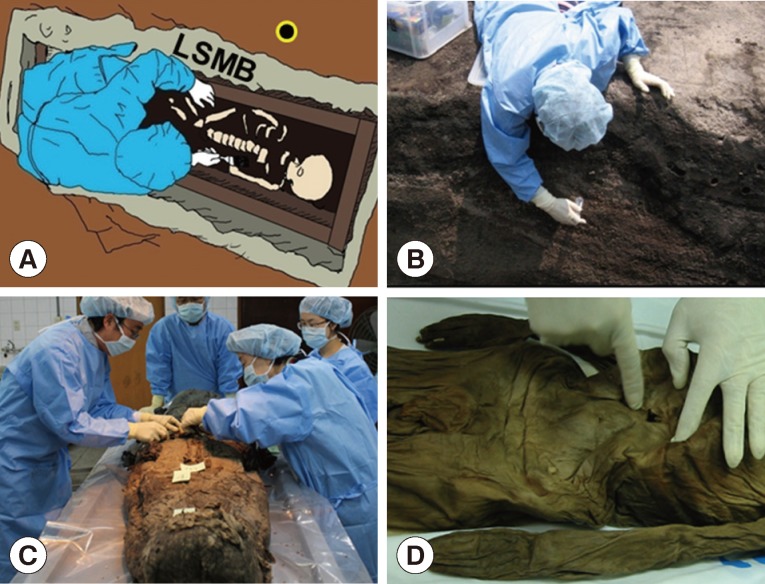
Fig. 3
Microscopic examination of the sample. Trichuris eggs (A) could be observed in coprolite sample of Yangju mummy. Bar=100 µm. Eggs of Trichuris trichiura (B) and Ascaris lumbricoides (C) in coprolite samples from Korean mummy. Bar=10 µm (B & C). (A) is from Yangju mummy, (B) from Mungeyong mummy, and (C) from SN1-2 case. Clonorchis sinensis egg (D) discovered in the geological strata sample. Since there were no parasite eggs in negative controls, the results could be regarded as authentic. Bar=10 µm. Scanning electron microscopic view of Trichuris trichiura (E) and Ascaris lumbricoides eggs (F) in the coprolite from a child mummy, Yangju. Bars=15 µm. 
Fig. 4
(A) A larva of Strongyloides stercoralis recovered from the Gongju mummy. (B) A larva of Trichostrongylus sp. recovered from the Gongju mummy. (C) An egg of Metagonimus yokogawai recovered from the Hadong-1 mummy. Bar=10 µm. (D) An egg of Gymnophalloides seoi recovered from the Hadong-1 mummy. Bar=10 µm. 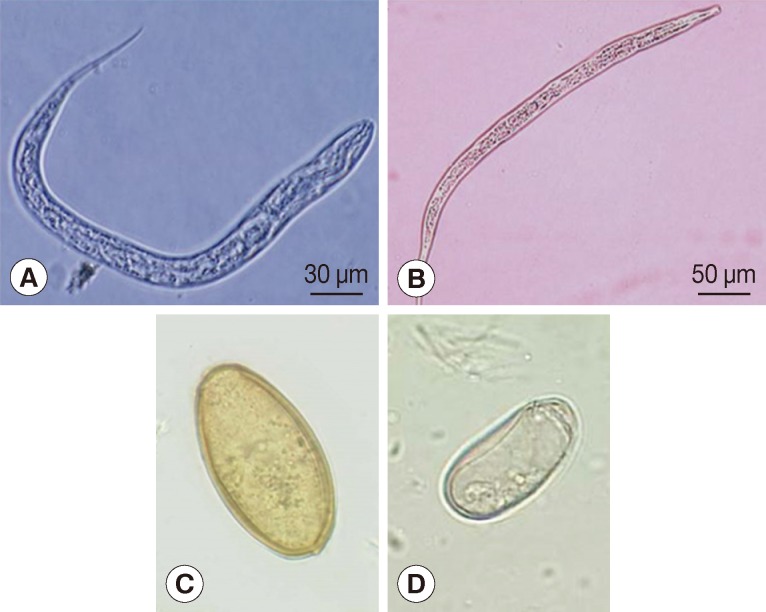
Fig. 5
Ectopic paragonimiasis in a Joseon female case. In the mummified liver sample, many Paragonimus westermani eggs were found. Bar=50 µm. 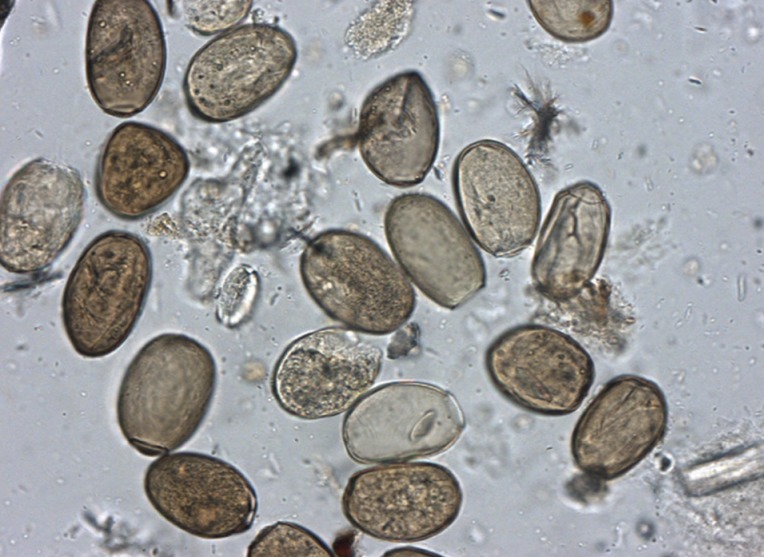
Fig. 6
(A) The places where 15 Korean mummies were discovered. Paleoparasitological studies have been done on the coprolites from each Korean mummy. 1, Yongin; 2, Jinju; 3, Sapgyo; 4, Hadong-2; 5, Hadong-1; 6, Sacheon; 7, Gangneung; 8, Dangjin; 9, Mungyeong; 10, Waegwan, 11, PJ-SM; 12, Seocheon; 13, Yangju; 14, SN (Sinnae); 15, GJ (Gongju). (B) Gymnophalloides seoi infection was identified in 2 cases of Korean mummies from (1) Sapgyo and (2) Hadong (indicated by blue dots). In a 2001 national survey, G. seoi infection was confined to small islands or counties in the southwestern corner of the Korean peninsula (areas shaded in red). 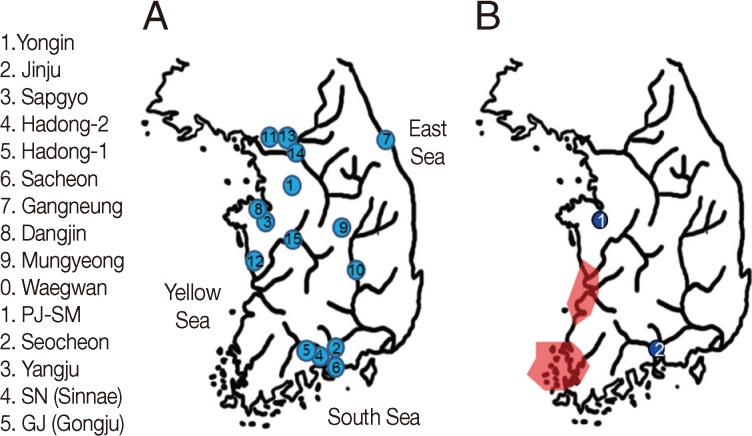
Table 1.
Parasite eggs discovered in mummies or ancient remains of Korea according to geographical areas
|
No. |
Area |
Estimated date |
Date of excavation |
Sex |
Parasite eggs from ancient mummies
|
|
Al |
Tt |
Hk |
Ev |
Cs |
Pw |
My |
Gs |
Ss |
Tr |
|
1 |
Yongin |
15-16C |
2007-8 |
Female |
+ |
+ |
|
|
|
+ |
|
|
|
|
|
2 |
Jinju |
15-16C |
2010-7 |
Female |
+ |
+ |
|
|
|
+ |
|
|
|
|
|
3 |
Sapgyo |
16C |
2011-2 |
Male |
+ |
|
|
+ |
|
+ |
+ |
|
|
|
|
4 |
Hadong-2 |
16-17C |
2009-6 |
Female |
|
|
|
|
|
+ |
|
|
|
|
|
5 |
Hadong-1 |
17C |
2006-4 |
Female |
|
|
|
|
+ |
|
+ |
+ |
|
|
|
6 |
Sacheon |
1620-1630 |
2011-10 |
Female |
|
|
|
|
|
|
+ |
|
|
|
|
7 |
Gangneung |
1622 |
2007-10 |
Male |
|
+ |
|
|
|
|
|
|
|
|
|
8 |
Dangjin |
1633 |
2008-8 |
Male |
+ |
|
|
+ |
|
+ |
|
|
|
|
|
9 |
Mungyeong |
1647 |
2010-4 |
Female |
|
+ |
|
|
+ |
|
|
|
|
|
|
10 |
Waegwan |
1648 |
2008-10 |
Male |
|
+ |
|
|
+ |
|
|
|
|
|
|
11 |
PJ SM |
1699 |
2008-5 |
Female |
+ |
+ |
|
|
|
|
|
|
|
|
|
12 |
Seocheon |
17C |
2007-2 |
Female |
+ |
+ |
|
|
|
|
|
|
|
|
|
13 |
Yangju |
1640-1700 |
2001-11 |
Male |
+ |
+ |
|
|
+ |
|
|
|
|
|
|
14 |
SN2-19-1 |
1765 ±10 |
2007-8 |
Male |
+ |
+ |
|
|
|
|
|
|
|
|
|
15 |
GJ1-2 |
17-18C |
2008-4 |
Male |
+ |
+ |
|
|
|
+ |
|
|
+ |
+ |
|
16 |
SN 1-2a
|
1605-1733 |
2007-8 |
Undecidable |
|
+ |
|
|
|
|
|
|
|
|
|
17 |
SN3-7-1a
|
16-17C |
2007-9 |
Undecidable |
|
+ |
|
|
|
|
|
|
|
|
|
18 |
SN2-19-2a
|
1755±10 |
2007-8 |
Undecidable |
+ |
+ |
|
|
|
|
|
|
|
|
|
Total 18 samples of mummies (15) & ancient remains (3); 9 (Al), 14 (Tt) 0 (Hk), 1 (Ev), 5 (Cs), 5 (Pw), 3 (My), 2 (Gs), 1 (Ss), 1 (Tr) |
Table 2.
DNA sequences of ancient parasite eggs from No. 12 (A. lumbricoides), No. 18 (T. trichiura), No. 4 (P. westermani), and No. 10 (C. sinensis) mummies
|
Species |
Target region |
Consensus sequence (5ʹ→3ʹ) |
Ref. |
|
Ascaris lumbricoides
|
18sRNA |
ATGGCCTATAAAGGTGAAACCGCGAACGGCTCATTACAACAGCTATTATATACTTGATCTTGATATCCTACGTGGATAACTGTGGTAATTCTAGAGCTAATACATGCACCAAAGCTCCGATTTTCTGACGAGCGCATCTATTAGATTAAAACCAATCGGGTTTCGGCCCGTCAATT |
[23] |
|
cyt b |
TTTTGACTTATGGTTGGAATTTTGGTAGTATGTTGGGTATGGTTTTAGGTTTTCAGATTTTGACTGGTACTTTTTTGGCTTTTTATTATTCTAATGAT |
|
|
Trichuris trichiura
|
SSUrRNA |
GGAACGATCGAGCCGTACATAAGTTCGGCTAAATCTATTGGAGGGCAAGTCTGGTGCCAGCAGCCGCGGTAATTCCAGCTCCAATAGCGTATATTAAAGTTGCTGCGGTTAAACCGCTCGTAGTTGGATTGCGGATGTCGACGACGGTCGTCCTAAGCAGGAGTCGTTCCGTCGCTCGTCACCTGTTCGATCAAGATTGCCCTGGATGCTCTTCAGTGAGTGTCCTTGGCGACTTGAAAGTTTACTTTGAGAAAATGAGAGCGCTCAAGGCAAGCCGTAGT |
[24] |
|
Paragonimus westermani
|
ITS2 |
ATATTGCGGCCACGGGTTAGCCTGTGGCCACGCCTGTCCGAGGGTCGGCTTATAAACTATCGCGACGCCCAAAAAGTCGCGGCTTGGGTTTTGCCAGCTGGCGTGATCTCCCCAATCTGGTCTTGTGCCTGTGGGGTGCCAGATCTGTGGCGTTTCCCTAACATACTCGGGCGCACCCACGTTGCGGCTGAAAGCCTTGACGGGGATGTGGCAACGGAATCGTGGCTCAGTGAATGATTTATGTGCGCGTTCCGCTGTCCTGTCTTCATCTGTGGTTTATGTTGCGCGTGGTCTGCTTTCGATGCTGACCTACGTATGTGCCATGTGGTTCATTCTCCTGACCTCGGATCAGACGTGAGTACC |
[40] |
|
COI |
Fail |
|
|
Clonorchis sinensis
|
ITS1 |
GTCTTGCAGCATTGTCTGCCTAGGGCGGAGCGATCCTAGTTCCGTCATGTTCTACATGTATGTTCCGCATGCATGCTGCAGCGTTGTCTGCCTACGGTGGAGCGTCTCTAGTTCTACCAATTCCTGGCTATACCTGGCACGTGTACCCAATATATATGATGTGCCTACGTACAGTCGCGTTTCGGCAGGGTGCCTACCCGTCTGATGCTCTCGGTATGCTCGCTTCCGTTGGTGGCCAGTCCATATTGGGGGTGACGGGATGTGCTGTCAGAATGGACAGTGCTAGGCTTAATGAGTGGGCATGATGTGTCTCGAGCTACGGCTCACCCACCGCCCTGATGTTGTTGTTCATTTCAAACCGTTTTACACTGTTAAAGTGTTTCAGGTTGGCCTGGCCTGGCCTGACTGGCTGGCCGGCTTGTCTCACTGCCCCGACATGCACCCGGTGTTCTACACTGGACTGCATGTGCAGTCGCCCGGCGGTGCCTTATC |
[39] |
|
ITS2 |
TGTCGATGAAGAGCGCAGCCAACTGTGTGAATTAATGCGAACTGCATACTGCTTTGAACATCGACATCTTGAACGCATATTGCGGCCATGGGTTTGCCTGTGGCCACGCCTGTCCGAGGGTCGGCTTATAAACTATCACGACGCCCAAAAAGTCGTGGCTTGGGTCTTGCCAGCTGGCATGATTTCCCCACACAATTGTGTGTATGTGTGTGGGGTGCCGGATCTATGGCTTTTCCCCAATGTGCCGGACGCAACCATGTCTGGGCTGACTGCCTAGATGAGGGGGTGGCGGCGGAGTCGTGGCTCAATTGTTGTTATTGTTGTGAATGTGCGCGCTCCGTTGTTGGTCCTTTGTCTTTGGTTGAGGCTTCAGTATTGGCAATGCATTCGATGCAAATCTGTTTTGCACCGGTCGGTGCTTAACTTTCCTGACCTCGGATCAGACGTGATT |
|
|
COI |
TATGATTAGTCACATTTGTACTACTTTAACAGGTAAAGATTCGTTGTTTGGTTATGGGGGCTTGGTGTTGGCTATGTTTGCTATAGTTTGTCTGGGTAGGGTGGTTTGAGCTCATCATATGTTTACTGTTGGGCTGGATTTGGGGACTGCTGTTTTTTTTAGCT |
|
|
|






