INTRODUCTION
Acarapis mites are parasites of bees which can cause severe damage to the bee industry by decreasing honey production, destroying colonies in both managed and feral honey bees and decreasing pollination [1]. Currently there are 3 species of Acarapis mites: A. woodi, A. externus, and A. dorsalis [2]. These mites are known to parasitize specific locations on bees. A. externus is mainly found on the external surface of the head and thorax of bees. A. dorsalis, on the other hand, is mainly found on the thorax especially in a groove between the mesoscutum and mesoscutellum of bees. Unlike these 2 species, A. woodi is an internal parasite that lives in the abdominal and thoracic air sacs of bees [3]. All stages of the mite live within the tracheae, except during a brief period when adult females disperse to search for new hosts [3].
Each Acarapis mite shows different pathogenicity according to its habitat. Although A. externus has been reported to cause wing loss or malfunction, the 2 species of external mites, A. externus and A. dorsalis, generally do little damages [1]. Unlike the other 2 ectoparasitic mites, A. woodi causes blockage of thoracic tracheae, reducing the diffusion of oxygen to the flight muscles and to the brain, and serve as a vector of other pathogens such as bacteria and viruses [3]. Moreover, heavy infestation of the mite causes a shortened lifespan of bees, a diminished brood [4], smaller bee populations, looser winter clusters, increased honey consumption, lower honey yields, and ultimate colony demise [3].
All 3 species of the genus Acarapis are distributed throughout many countries. A. dorsalis has been reported on Apis mellifera from Europe, Canada, USA, New Zealand, Australia, Papua New Guinea, and Hawaii. The world distribution of A. externus is the similar to that of A. dorsalis, but A. externus is collected more frequently and more abundant than A. dorsalis [2]. A. woodi has been reported in many European countries, including Scotland, France, Spain, and Greece, after its first report in 1919. The mite then spread to the American continent, first to Argentina, Colombia, Mexico, and USA. Outside the European and American continents, it was reported in Iran, Turkey, China, and Japan [2,5-7]. Because Korea is located between China and Japan, Acarapis mites are expected to be present in Korea. However, the presence of A. woodi in Korea has not yet been reported.
In Korea, many studies on parasitic bee mites have been reported, including the infestation status and control methods on Varroa jacobsoni and Tropilaelaps clareae [8]. However, there have not yet been studies on their occurrence. If an outbreak of the mites suddenly happened, this present situation could cause bee colony devastation and Acarapis mite-mediated diseases on the Korean apicultural industry. We used a PCR detection method specific to Acarapis mites and surveyed the prevalence of Acarapis mites.
MATERIALS AND METHODS
Sample collection
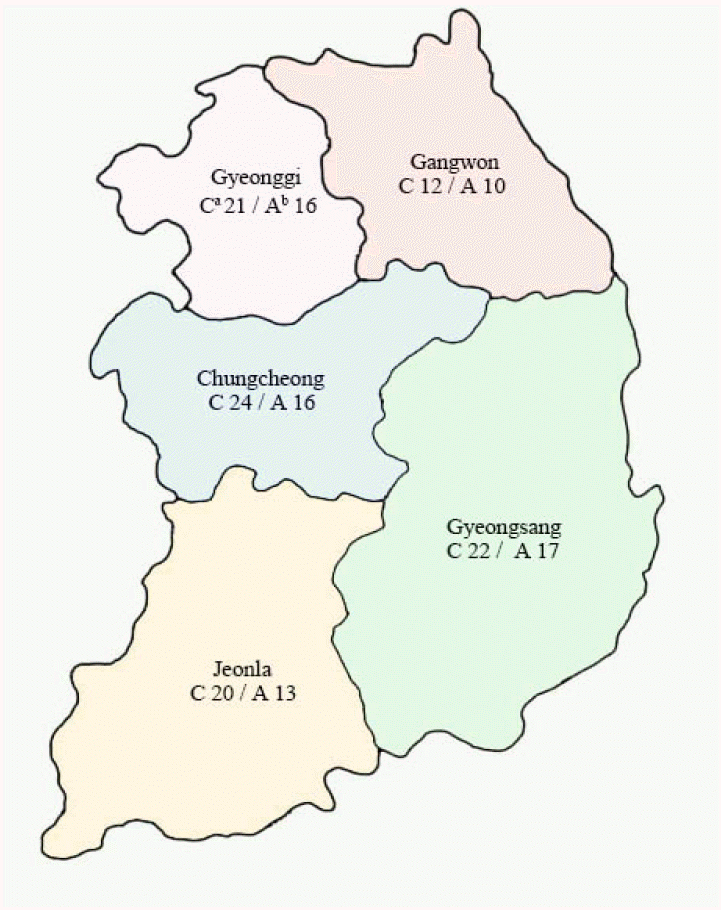
A total of 99 Apis mellifera colonies were sampled from 72 apiaries in 5 provinces (Gyeonggi, Gangwon, Chungcheong, Gyeongsang, and Jeolla) in Korea (Fig. 1). Fifty to 100 adult bees were collected and pooled from 1-2 colonies in each apiary between August and December 2013. The honey bee samples were collected from brood nests inside the hives for managed colonies and at the hive entrances for feral colonies, respectively [5]. After collecting the bees, the samples were stored at -20˚C for PCR detection.
PCR detection and sequencing
The head and thorax of 20 adult bees randomly chosen from each hive were used for DNA extraction using DNeasy Blood & Tissue kit (Qiagen, Hilden, Germany) following the manufacturer’s instructions. Total DNA was eluted in 30 µl of elution buffer and used for PCR with LeGene Hot Start PCR Mix w/Sky Blue Dye (LeGene Biosciences, San Diego, California, USA). Two primer sets, including Acarapis (A) primer and Kojima (K) primer set, were used to identify Acarapis mite (Table 1) which were capable of amplifying mitochondrial cytochrome c oxidase subunit I (COI) DNA fragments of Acarapis mites [5,9,10]. These primers do not amplify honey bee COI DNA fragments; thus, total DNA isolated from bees was used as a template for PCR. As a positive control for PCR to verify the quality of DNA extraction, a honey bee genomic DNA fragment encoding a part of a honey bee Hymenoptera-specific transient receptor potential A channel (AmHsTRPA) was amplified by AmHsTRPA (T) primer set [10]. The thermal cycling conditions were as follows: 1 cycle of initial denaturation at 94˚C for 2 min, 35 cycles of denaturation at 94˚C for 30 sec, annealing at 55˚C for 30 sec, and extension at 72˚C for 30 sec. The PCR product was analyzed by 2% agarose gel electrophoresis. A negative control lacking template DNA was performed for each PCR reaction. Since A and K primer sets equally amplify COI genes of 3 species of Acarapis mites (A. woodi, A. dorsalis, and A. externus), the amplified products with T and A primer sets or T and K primer sets were sequenced and aligned with the sequences deposited in GenBank to determine the parasitized mite species. The nucleotide sequences were identified by the Basic Local Alignment Search Tool (BLAST) at the National Center for Biotechnology Information (NCBI). Multiple nucleotide alignment was carried out using the published SBV sequences as references by BioEdit version 7.0.9.0 (Table 2). The phylogenetic tree was constructed by Mega 6.06.1 software [11] using the neighbor-joining (NJ) method [6].
Microscopic examination of Acarapis mite
To confirm the presence of Acarapis mites, a total of 20 bees per each colony were examined with a modified dissection method developed by Lorenzen and Gary [12]. The external surface, including the head, thorax and a groove between the mesoscutum and mesoscutellum of each bee, was examined to find out A. externus and A. dorsalis. To examine A. woodi, each bee was pinned through the thorax into bee wax culture dish and placed under a dissecting microscope. The head and the first pair of legs were removed off using a scalpel or razor blade. The ring of prothoracic sclerite (collar) was also removed using fine forceps to fully expose both left and right sides of the trachea. The exposed trachea of 2 right and left sides was examined with 20× magnification to find out tracheal color change, the body of A. woodi and foreign substance. If the presence of A. woodi or foreign substance was confirmed, the trachea was detached and placed on a glass slide and examined under an optical microscope to diagnosis A. woodi.
RESULTS
Prevalence of Acarapis mite infestations
A total of 99 colonies from 72 apiaries in 5 provinces (Gyeonggi, Gangwon, Chungcheong, Gyeongsang, and Jeolla) were examined (Fig. 1). All of the 99 colonies were T primer set positive. Among these, 36, 2, and 4 colonies were positive for A, K, and both A and K primer sets, respectively (Table 2). The infection rate of Acarapis mites in Gangwon, Gyeonggi, Chungcheong, Gyeongsang, and Jeolla were 25.0%, 9.5%, 25.0%, 95.5%, and 50.0%, respectively (Table 2). Gyeongsang had the highest infection rate, followed by Jeolla, Chungcheong, and Gyeonggi. Gangwon had the lowest infection rate among the 5 provinces. Between the primer sets, the A primer set was more sensitive than the K primer set (Table 2).
Molecular characterization of Korean Acarapis mites
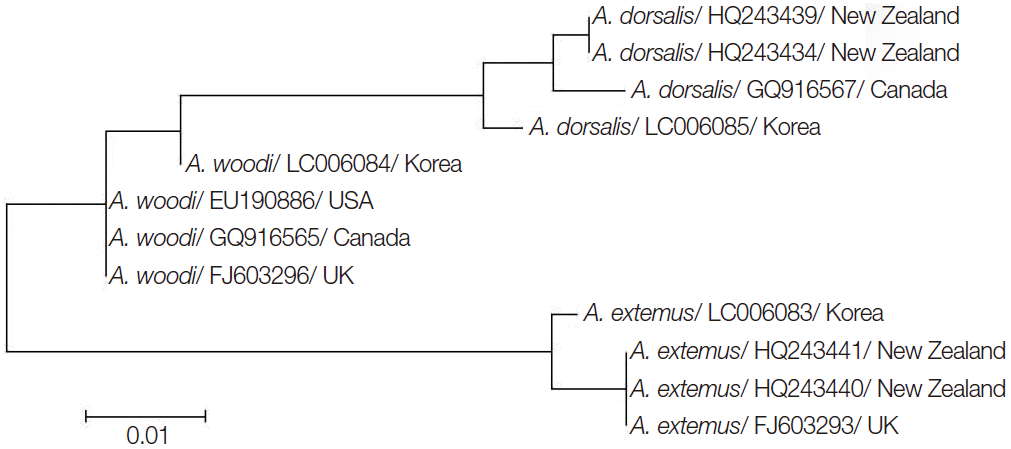
To confirm the above PCR assay results, all 42 Acarapis-positive colonies were sequenced and aligned. Each sequence showed 92.6-99.3% homology with reference sequences available in GenBank (Table 3). Species identification of each colony was determined if the sequence between the unknown sample and the reference showed 98-100% homology. Based on these criteria, the number of colonies infected with A. dorsalis was 32 samples which showed the highest infection rate among the 3 species, while the number of colonies infected with A. externus and A. woodi was 9 and 1, respectively. The DNA sequences of PCR-amplified CO1 fragments infested by A. dorsalis A. externus, and A. woodi were deposited to a DDBJ database as the no. LC006085, LC006083, and LC006084, respectively. Although A. externus and A. dorsalis were distributed throughout all 5 provinces, A. woodi was only detected from an apiary in Mungyung City, Gyeongsang (Gyeongsangbuk-do Province) (Table 4). The phylogenetic analysis between Korean isolates in this study and other references was performed (Fig. 2). Each Korean isolate Acarapis species was placed on a separate branch and closely related to the same Acarapis species.
Morphological diagnosis of Acarapis mites
In microscopic examinations, a total of 20 bees per each colony were dissected, but no diagnosable mite body was detected. However, foreign bodies which assumed the mite and some debris were discovered from the A. woodi-positive bee colony (Fig. 3).
DISCUSSION
In this study, the prevalence of Acarapis mites was surveyed by PCR and subsequent sequencing. Molecular analysis of bee DNA extracts using PCR and alignment indicate that all 3 species of Acarapis mites are present in Korea and 32.3% (32/99) colonies were infected with A. dorsalis, 9.1% (9/99) with A. externus, and 1.0% (1/99) with A. woodi from 99 colonies in Korea. Although we molecularly identified Acarapis mites, the Acarapis mites were not found in microscopic examinations. This result could be explained by the fact that all apiaries in the survey used in-house type acaricides against bee mites such as Varroa destructor and Tropilaelaps clareae which also affected against Acarapis spp. Reports also indicate that most apiaries of Korea regularly use insecticides for acarine control [13,14]. Therefore, only dead bodies of the mites or debris might have remained and been detected by PCR assay (Fig. 3).
While A. dorsalis and A. externus were detected in all 5 provinces, A. woodi was found only in 1 apiary farm, and the infection rate was very low. Species-specific seasonal fluctuation could explain this phenomenon. In A. dorsalis and A. externus, the incidence of these mites decreased in fall, increases in spring, and is highest during May and September in temperate regions [15,16]. On the other hand, the prevalence of A. woodi Table 3. increased greatly from November to February and declined in late spring [17]. We collected most of the bee samples during August and October. Consequently, the population of A. woodi might be too low for detection. In future studies, the bee samples should be collected during fall and winter seasons and examined to confirm the seasonal fluctuation of Acarapis mites in Korea.
In general, the most common diagnostic method for Acarapis mite infestation is morphological identification. Further, the molecular prevalence of Acarapis mites has been conducted only in a few countries [14]. However, the morphological identification is generally time-consuming and requires detailed and sustained attention by the screener. On the other hand, molecular methods facilitate rapid and sensitive detection of Acarapis spp. in many honey bee samples for epidemiologic surveys [10]. Therefore, we adopted 2 different genusspecific primers, including K and A primer sets. Our results indicated that the A primer set was more sensitive than the K primer set. However, both A and K primer sets require sequencing of amplicons to identify the species of the mite. Therefore, there is a need to design a primer which can amplify species-specific COI DNA fragments in future studies.