Warning: mkdir(): Permission denied in /home/virtual/lib/view_data.php on line 81
Warning: fopen(upload/ip_log/ip_log_2024-04.txt): failed to open stream: No such file or directory in /home/virtual/lib/view_data.php on line 83
Warning: fwrite() expects parameter 1 to be resource, boolean given in /home/virtual/lib/view_data.php on line 84 Antibody-dependent rat macrophage-mediated damage to the excysted metacercariae of Paragonimus westermani in vitro
Antibody-dependent rat macrophage-mediated damage to the excysted metacercariae of Paragonimus westermani in vitro
P R Chung,J K Chang and C T Soh*
Department of Parasitology, College of Medicine, Yonsei University, Seoul, Korea.
Abstract
An in vitro immune effector mechanism against the target excysted metacercariae of Paragonimus westermani was demonstrated in the rat system. Peritoneal exudate cells, mainly macrophages from normal rats, showed adherence to and killing of excysted metacercariae of P. westermani in the presence of complement-independent serum from rats infected with Paragonimus metacercariae. These reactions were specific for the excysted metacercariae, as tissue-migrating juvenile worms were not affected.
Damage of excysted metacercariae of P. westermani due to antibody and macrophages was assessed by morphological observation, by cell adherence reaction and by the use of vital dyes. Trypan blue dye exclusion proved to be a reliable indicator of judging metacercarial viability. Electron microscopic studies demonstrated that macrophages reacted with fuzzy material on the tegumental surface and fine structures in the syncytium of the parasites. The tubular tunnels formed between the basement membrane and muscle layers of the damaged parasites were also noticeable. The relevance of these findings to cellular immunity in the early paragonimiasis was discussed.
Figures
Fig. 1 Antibody-dependent adherence of rat peritoneal macrophages to the excysted metacercariae of P. westermani. (A) in the presence of macrophages and sera from uninfected normal rats, the excysted metacercariae appear normal and cells do not adhere to worms; (B) I the presence of sera from infected rat and macrophages, cells adhere to the surface of worms; (C) blebbing of the tegument with strong cell adherence reaction appears on the surface of dead worm, in the presence of sera from the rats 2 weeks after infection. (×100).
Figs. 2-4 Fig. 2. An excysted metacercaria of P. westermani after incubation in media containing sera from uninfected rats shows the normal, intact structural features such as tegumental ridges (TR), tegumental syncytiun (TS) containing mitochondria (Mi) and small ovoidal dense granules, circular muscles (CM), and longitudinal muscle (LM) layers just underneath the basement membrane (BM) of the tegumental syncytium (×12,500).
Fig. 3. Appearance of an excysted metacercaria after 2-hr incubation in media containing macrophages (Mac) from uninfected and sera from infected rats. Note deposition of electron-dense fuzzy material (FM) onto the tegument. Syncytial cytoplasm contains spines (Sp) and intact dense granules (arrow heads) (×10,000).
Fig. 4. Antibody-dependent adherence of macrophages and damage to the excysted worm. The excysted metacercariae were incubated for 18 hrs in the presence of macrophages (Mac) from uninfected rats and serum from infected rats. Note macrophages with a great number of pseudopodia (Ps) onto the tegumental surface and dense granules surrounded by small vesicles in the syncytial cytoplasm (arrow heads). No tegumental ridges were observed (×7,500).
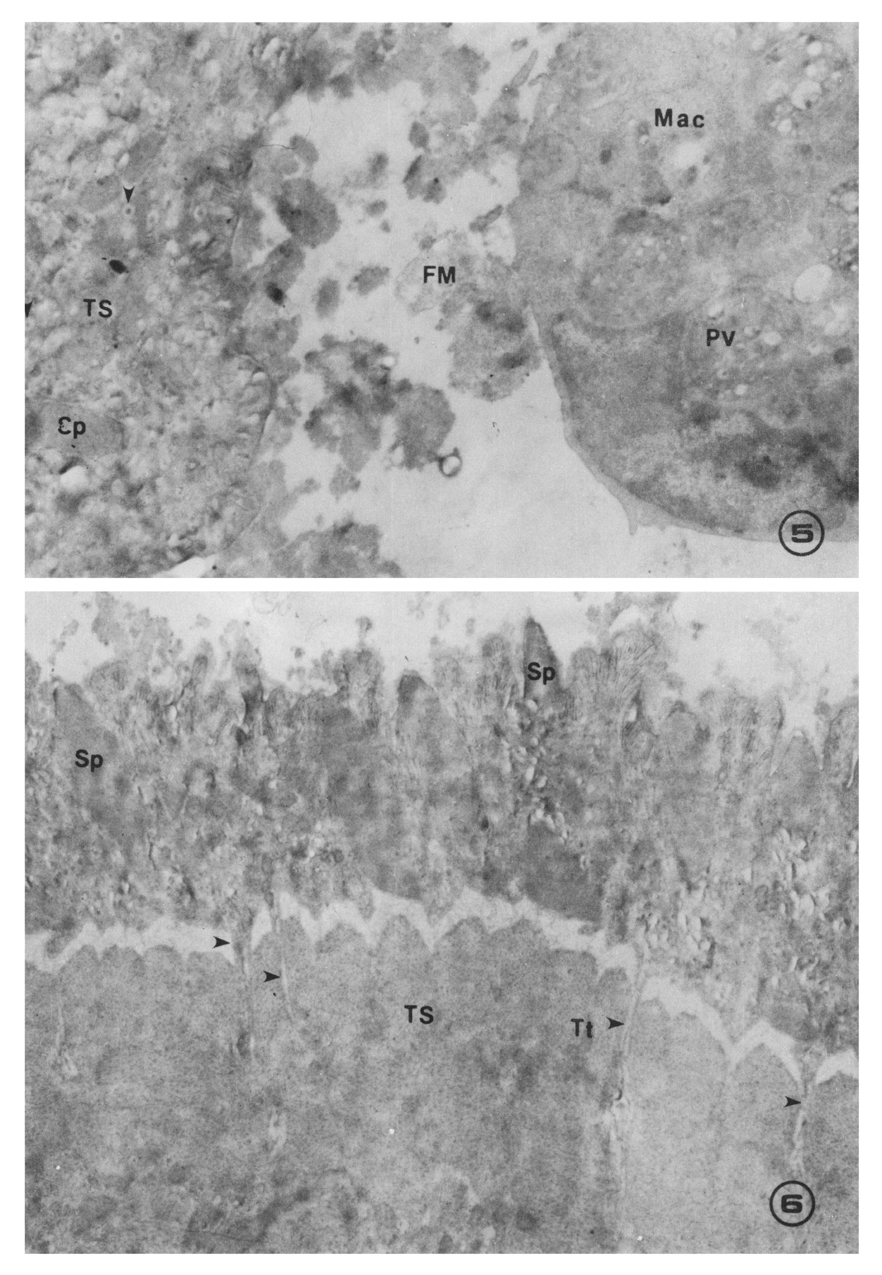
Figs. 5-6 Fig. 5. An excysted metacercaria and a macrophage incubated in the same effector system as in Fig. 4. The increased amount of fuzzy material (FM) between macrophage and tegumental surface of worm, and phagosome-like vacuoles (PV) in the macrophage were remarkable. Dense granules denatured in the small vesicles were also enlarged (×12,500).
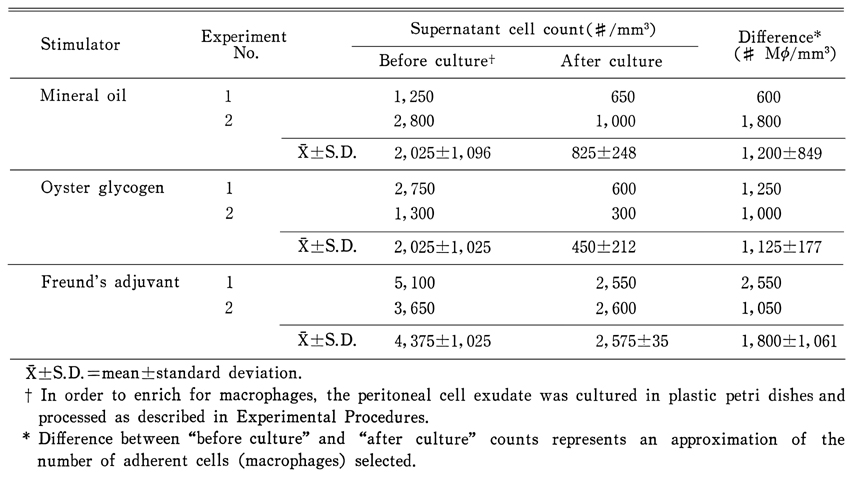
Fig. 6. A dead worm incubated form 18 hrs in the presence of homologous macrophages and serum obtained from rats 2 weeks after infection. Tubular tunnels (Tt; arrow heads) formed between the basement membrane and muscle layers of a damaged parasite were noticeable. The fine structures in the tegumental syncytium were almost destroyed (×10,000).
Tables
Table 1 Mean worm recoveries from five rats infected with 50 Paragonimus metacercariae per animal*
Table 2 Yield of rat peritoneal macrophages (Mø) according to stimulators employed
Table 3 Cell adherence and death of excysted metacercariae of P. westermani after an 18-hour culture with immune sera collected from rats at one-week intervals*
Table 4 Cell adherence and viability of juvenile worms(=adolescariae) of P. westermani in various effector system*
References
1.
Butterworth AE, David JR, Franks D, Mahmoud AA, David PH, Sturrock RF, Houba V. Antibody-dependent eosinophil-mediated damage to 51Cr-labeled schistosomula of Schistosoma mansoni: damage by purieid eosinophils. J Exp Med 1977;145(1):136–150.
2.
Butterworth AE, Sturrock RF, Houba V, Taylor R. Schistosoma mansoni in baboons. Antibody-dependent cell-mediated damage to 51Cr-labelled schistosomula. Clin Exp Immunol 1976;25(1):95–102.
3.
Butterworth AE, Sturrock RF, Houba V, Mahmoud AA, Sher A, Rees PH. Eosinophils as mediators of antibody-dependent damage to schistosomula. Nature 1975;256(5520):727–729.
4.
Capron A, Dessaint JP, Capron M, Bazin H. Specific IgE antibodies in immune adherence of normal macrophages to Schistosoma mansoni schistosomules. Nature 1975;253(5491):474–475.
5.
Chung PR. Yonsei Rep Trop Med 1983;14(1):44–56.
6.
Chung PR, Asch HL, Bruce JI. Antibody-dependent murine macrophage-mediated damage to Schistosoma mansoni schistosomula in vitro. Cell Immunol 1982;74(2):243–259.
7.
Dean DA, Wistar R, Chen P. Immune response of guinea pigs to Schistosoma mansoni. I. In vitro effects of antibody and neutrophils, eosinophils and macrophages on schistosomula. Am J Trop Med Hyg 1975;24(1):74–82.
8.
Dean DA, Wistar R, Murrell KD. Combined in vitro effects of rat antibody and neutrophilic leukocytes on schistosomula of Schistosoma mansoni. Am J Trop Med Hyg 1974;23(3):420–428.
9.
Kassis AI, Aikawa M, Mahmoud AF. Mouse antibody-dependent eosinophil and macrophage adherence and damage to schistosomula of Schistosoma mansoni. J Immunol 1979;122(2):398–405.
10.
Kum PN, Nchinda TC. Pulmonary paragonimiasis in Cameroon. Trans R Soc Trop Med Hyg 1982;76(6):768–772.
11.
Min DY, Ahn MH, Kim KM, Leem MH, Park SY. [The effects of antibodies and complement in macrophage-mediated cytotoxicity on metacercariae of the lung fluke, Paragonimus westermani]. Korean J Parasitol 1990;28(2):91–100.
12.
Miyazaki I. aa. Jpn J Parasitol 1974;23(5):280–284.
13.
Miyazaki I. J Parasitol 1967;53:207.
14.
Oshima T. aa. Jpn J Parasitol 1956;5:403–415.
15.
Perez HA, Smithers SR. Schistosoma mansoni in the rat: the adherence of macrophages to schistosomula in vitro after sensitization with immune serum. Int J Parasitol 1977;7(4):315–320.
16.
Ramalho-Pinto FJ, McLaren DJ, Smithers SR. Complement-mediated killing of schistosomula of Schistosoma mansoni by rat eosinophils in vitro. J Exp Med 1978;147(1):147–156.
17.
Sher A. Complement-dependent adherence of mast cells to schistosomula. Nature 1976;263(5575):334–336.
18.
Smithers SR, Terry RJ. The immunology of schistosomiasis. Adv Parasitol 1976;14:399–422.
19.
Soh CT. aa. Yonsei Med J 1962;3(1):79–84.
20.
Walton BC, Chyu I. Clonorchiasis and paragonimiasis in the Republic of Korea. Report on a prevalence survey using intradermal tests. Bull World Health Organ 1959;21:721–726.
21.
Yong TS, Kim TS, Lee JS, Lee OY, Kim DC. Detection of circulating antigens in rats experimentally infected with Paragonimus westermani by ELISA. Korean J Parasitol 1987;25(2):141–148.